F1 Crosses of Evergreen and Deciduous Azaleas
and Other Wide Crosses of Rhododendron
Richard A. Jaynes, New Haven, Connecticut Associate Geneticist,
The Connecticut Agricultural Experiment Station, New Haven
There is a dearth of experimental information on the natural affinities among Rhododendron species in spite of the fact that a large number of species hybrids were produced in the past one hundred years. This has come about partly because most hybridization efforts were directed at developing improved ornamentals. Pure species were often avoided as parents and hybrids of unknown pedigree were used in their stead.
If we take a simplistic approach, and assume the classification of the species is correct (Leach 1961, Wilson and Rehder 1921), we would expect crosses between Rhododendron series to be less likely to succeed than crosses among more closely related species within series. This does not seem to be the case. Some of the first Rhododendron hybrids observed, called azaleodendrons, were natural crosses between azaleas (subgenus Anthodendron) and other Rhododendron series. Yet, some of the most difficult crosses to complete successfully are crosses within the azalea series. A thorough understanding of crossability among all the species of Rhododendron will probably never be achieved but surely more will be learned about the mechanisms of compatibility. With this information will come a better understanding of the genetic affinities and rationale for the patterns of crossability observed.
The 74 azalea species recognized by Leach comprise only one of the 43 series within the genus Rhododendron and are further divided into six subseries. Numerous azalea hybrids and cultivars have been developed by breeders but most azalea hybridization has been confined to species within series.
Pryor (1973) briefly reviewed the literature on evergreen and deciduous azalea crosses and also reported several crosses that resulted from an extensive USDA breeding program. The successful USDA crosses were largely the result of
R. kaempferi
or
R. indicum
hybrids crossed with
R. mollis
cultivars.
Martin (1970), attempted approximately 1,000 azaleodendron crosses but obtained viable seedlings from only a few crosses. He was most successful using the azaleas as pollen parent. He succeeded in crossing both deciduous and evergreen azaleas to rhododendrons in other series including at least one cross of a lepidote (
R. carolinianum
) with an elepidote (
R. mollis
). A list of the crosses was not given, apparently few were between evergreen and deciduous azaleas.
Wada (1974) reported several putatively successful crosses of evergreen azaleas with other rhododendrons including
R. racemosum
,
R. keiskei
,
R. yakushimanum
, and
R. degronianum
. In addition he stated that evergreen azaleas (Obtusum subseries) hybridize rather freely with a number of the trifoliate deciduous azaleas in Schlippenbachii subseries; however, documentation of many of these crosses is needed.
Other wide crosses are known such as the putative hybrid of
R. roseum
and
R. carolinianum
produced by Dorothy Knippenberg and named 'Carolina Rose'. August E. Kehr (personal communication) has crossed
R. mucronulatum
x
R. obtusum
'Takei' and also
R. mucronulatum-chapmanii
x
R. mollis
; in both cases the plants were sterile and have since been lost.
The pattern in general with azaleas is that crosses between species belonging to different subseries within the series almost invariably fail to yield viable hybrids. The literature refers to such failures but little documentation of specific crosses is available. Generally only the successful crosses are reported and most often only those yielding plants of possible horticultural merit. If we accept 74 species of azaleas then there are 5,402 interspecific combinations possible within the series, including reciprocals. To a breeder there is always the nagging question of exactly which species have been crossed, for perhaps one of those not tested will prove to be a valuable genetic bridge between previously isolated species.
The purpose of this study was to develop a better understanding of the genetic barriers to interspecific gene exchange among azaleas in different subseries. For various reasons progress in this direction was limited but a summary of the results of Ft crosses among the azalea species is presented for its possible value to systematists and other breeders.
Materials and Methods
Plants were obtained from many commercial sources, from the U.S. Plant Introduction Station, as well as from collections in the wild. Since the genetic barriers between species were of prime interest, wild collected material was preferred, although not used exclusively. Crosses were repeated using different clones and in reciprocal combination whenever possible.
Flowers were emasculated in the bud stage and pollinated the following day. Stored (frozen) pollen was used when the flowering periods of the parent species did not overlap. A record was kept of the parent plants, date of cross, number of flowers, whether pollen was fresh or stored, capsule set, and seed set. If seed was obtained it was sown in an artificial soil mix in plastic boxes and kept under fluorescent lights and observations recorded on germination, chlorotic seedlings, and subsequent growth. Surviving seedlings were grown on in the greenhouse and field. Results and Discussion Results and observations are summarized in the accompanying chart. Other species used less extensively included
R. bakeri
,
R. reticulatum
, and
R. mucronatum
. Results of crosses between subseries with these few additional species fit the usual pattern and yielded no viable seedlings.
Generally no seed was produced from crosses between subseries. This was particularly true from the crosses of species in Schlippenbachii subseries or Canadense subseries. Notable, however, was the seed set and, in a few cases, viable seedlings from some crosses of species in Luteum subseries with those in Obtusum subseries. Of the nine such combinations that produce viable seedlings, the five with near normal appearing seedlings were
R. alabamense
x
R. obtusum
(including reciprocal),
R. oblongifolium
x
R. obtusum
,
R. oblongifolium
x
R. kaempferi
,
R. oblongifolium
x
R. poukhanense
(including reciprocal), and the pictured cross of
R. viscosum
x
R. poukhanense
1
.

|
|
Rhododendron viscosum
x
R. poukhanense
. The flowering period is
between that of the parental species. The hairy leaves and the lavender flower color are characteristic of the pollen parent (Scale in cm). Photo by Richard Jaynes |
Flowers were obtained on one or more seedlings from these five crosses, except for
R. alabamense
x
R. obtusum
, and all were pollen and seed sterile. The seedlings were intermediate in leaf size, shape, and hairiness, as well as flowering time, all tended to retain a few leaves just below the terminal bud. The effect of the pollen parent in modifying traits of the seedlings was clear in all cases and thus the success of the crosses verified. The tendency for all the hybrids to be evergreen is in contradiction to the observations of Pryor (1974) who reported leaf retention to be positively correlated with the maternal parent. The reason for this difference in results is not clear but may be related to the species used.
Reciprocal differences were noted for some crosses but in general over-all success or failure of a cross was not dependent on the direction of the cross. An example of a reciprocal difference observed was that
R. nudiflorum
x
R. kaempferi
produced no seed whereas the reciprocal cross produced seed that germinated poorly, yielding yellow green inevitable seedlings.
A few wide crosses were attempted among the azaleas and other rhododendrons as well as with
Kalmia latifolia
. The only successful cross was that of
R. oblongifolium
(elepidote) with
R. mucronulatum
(lepidote) which produced viable but sterile seedlings. Large scales were not present on the seedlings but were partially replaced with small stalked glands, an observation not unlike that reported by Heyting (1970) for another elepidote x lepidote cross. Based on this cross as well as the results of the Luteum and Obtusum azalea crosses,
R. oblongifolium
is a comparatively good bridging species.
Self-pollination of most of the species tested did not result in seed. Seedlings grown from a selfed
R. nudiflorum
plant were weak and spindly, symptoms characteristic of inbreeding depression.
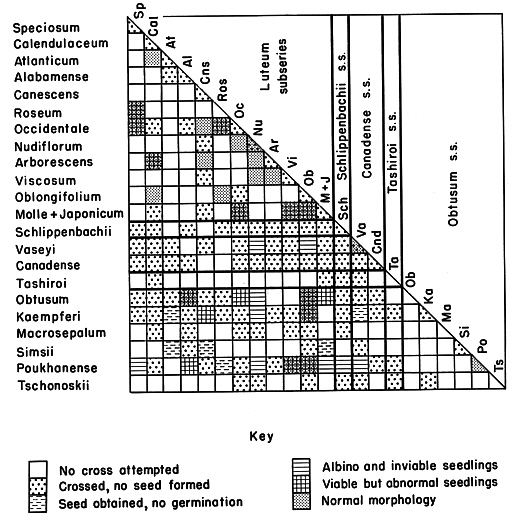
|
|
Interspecific crosses among several azalea species. (Crosses of different parent plants
of the same species occasionally give different results as indicated in the chart.) |
Summary
Seed set, germination, and even viable seedlings, were obtained from crosses of evergreen with deciduous species. The seedlings were morphologically intermediate to the parents. However, the F' hybrids were sterile and judged of little value in future breeding. The majority of more than 125 crosses produced no seed. In general, the results reinforce the conclusions previously drawn in the literature as to the rather substantial genetic barriers between azalea species belonging to different subseries.
Herbarium specimens of these five interspecific crosses are available at the Connecticut Agricultural Experiment Station. The taking of such pressed, documentary specimens is a common practice with botanists and taxonomists. It should become a more common practice among hybridizers and is especially useful with plants of unique or questionable parentage.
Literature Cited
1) Heyting, J., 1970, Hybrids between Elepidote and Lepidote Rhododendrons, Quart. Bull. A.R.S., 24:97-98.
2) Leach, D. G., 1961, Rhododendrons of the World, Charles Scribner's Sons, N.Y. 544 pp.
3) Martin, A. C., 1970, Breeding Azaleodendrons, Quart. Bull. A.R.S., 24: 39-41.
4) Pryor, R. L., 1973, Hybridization Between Evergreen and Deciduous Azaleas, Quart. Bull. A.R.S., 27:212-214.
5) Wada, K., 1974, Hybridizing Myths of Evergreen Azaleas, Quart. Bull. A.R.S., 28:101, 104-105.
6) Wilson, E. H. and A. Rehder, 1921, A monograph of azaleas (Rhododendron subgenus Anthodendron), Pub. Arnold Arb. No. 9, 219 pp.