INCUBATION TEMPERATURE AND POLLEN TUBE GROWTH IN RHODODENDRON:
A PRELIMINARY REPORT
Contribution No. 568, Kansas Agricultural Experiment Station
Robert J. Campbell, Assistant Professor of Floriculture
Department of Horticulture and Forestry, Kansas State University, Manhattan, KA
Interspecific
Rhododendron
crosses that consistently produce no seeds may fail because the pollen tubes do not reach the ovules. Stylar heat treatment might overcome this interspecific incompatibility for these reasons.
First, most
Rhododendron
species are self incompatible (7). This intra-specific self incompatibility in Rhododendron, not yet adequately described, resembles the gametophytic self-incompatibility system found in the genera
Lilium, Nemesia
and
Trifolium
(2,9,10). In these three genera and Rhododendron, pollen, when shed from the anthers, is binucleate (4) and germinates on the stigma and grows down the style in both self-incompatible and compatible pollinations (5,12), even though fertilization occurs only after compatible pollinations. Second, stylar heat treatment overcomes this gametophytic self incompatibility so that seed is set from self-incompatible pollinations. In the lily style, intra-specific compatible and self-incompatible pollen tube growth rates depend on temperature (2). At 22°C (72°F) during 48 hr of incubation, self-incompatible pollen tubes grow half the length of the 100 mm lily style while compatible pollen tubes grow almost through the style. At 34°C (95°F), however, self-incompatible pollen tubes grow to the same length as compatible pollen tubes at 22°C, long enough to cause self seed set. In various self-incompatible species including
Nemesia, Trifolium
and
Chrysanthemum
, incubation of blooming plants at about 34°C for 24 to 48 hr followed by self pollination resulted in good seed set (6,8,1 1).
In crosses between distantly-related
Rhododendron
species, high-temperature treatment of the style may or may not affect incompatible pollen tube growth. In
Lillium
(3) and in
Rhododendron
(5), interspecifically pollinated styles incubated 48 hr at 22°C exhibit two patterns of inadequate pollen tube growth: 1) Short-growth pollen tubes, grossly deformed, stopped just inside the stigma. 2) Half-growth pollen tubes resembling self-incompatible pollen tubes. Stylar heat treatment in
Lilium
does not affect short-growth pollen tubes resulting from interspecific pollinations. In some cases involving half growth pollen tubes, however, the stylar heat treatment increased pollen tube lengths so that the pollen tubes might have entered the ovary (1). Thus, if interspecifically incompatible pollen tubes in
Rhododendron
react to heat treatment as do half-growth, interspecific pollen tubes in
Lilium
, stylar heat treatment may produce seed set in crosses between
Rhododendron
species previously considered impossible to cross.
I am now studying the effects of incubation temperature on intra- and inter-specific pollen tube growth in Rhododendron. In June 1975, in Oregon, I cut just-opening flowers of several Rhododendron species and cultivars; placed the cut stems in vases of water; opened them in a growth chamber at 22°C; stripped petals and stamens from the flowers leaving the pistil, receptacle and a short pedicel piece; and then placed pollen either on its own styles, on styles of a closely related species or cultivar, or on styles of a distantly related species. I laid the pollinated styles on moistened filter paper in Petri plates and incubated them for 48 hr at 15, 20, 25, 30 or 35°C. I then submerged the styles in a killing solution of phenol: lactic acid: glycerin (1:1:1) and am storing them until I examine the styles for pollen tube growth using fluorescent microscopy. So far, I have cross-pollinated the evergreen elepidote rhododendrons, both as male and female, with the deciduous American azaleas, the evergreen lepidote rhododendrons and the evergreen azaleas. Also, I have reciprocally cross-pollinated the evergreen lepidote rhododendrons and the evergreen azaleas (See diagram). I plan to continue the work in Oregon in May and June 1976.
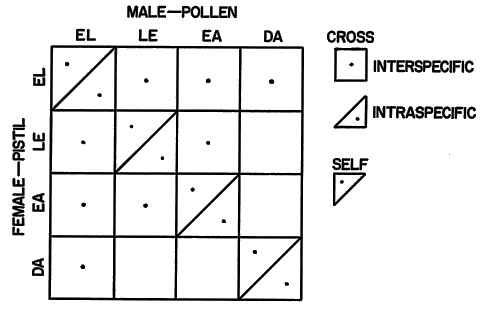
|
|
Azalea and rhododendron groups cross-pollinated during 1975 to study the
effect of incubation temperature on intra- and inter-specific pollen tube growth in Rhododendron. The four groups being studied are: Evergreen elepidote rhododendrons (EL), evergreen lepidote rhododendrons (LE), evergreen azaleas (EA) and deciduous azaleas (DA). |
REFERENCES
- Ascher, P. D., 1975, Personal communication.
- Ascher, P. D. and S. J. Peloquin, 1966, Influence of temperature on incompatible and compatible pollen tube growth in Lilium longiflorum, Can. Journ. Genet. Cytol., 8:661-664.
- Ascher, P. D. and S. J. Peloquin, 1968, Pollen tube growth and incompatibility following intra- and inter-specific pollinations in Lilium, Amer. J. Bot., 55:1230-1234.
- Brewbaker, J. L., 1967, The distribution and phylogenetic significance of binucleate and trinucleate pollen grains in the angiosperms, Amer. J. Bot., 54:1069-1083.
- Campbell, R. J., 1975, Unpublished data.
- Campbell, R. J. and P. D. Ascher, 1972, High temperature removal of self-incompatibility in Nemesia strumosa, Inc. Newsletter, 1:3-5.
- Cox, P. A., 1973, Dwarf Rhododendrons, Macmillan Publishing Co., New York, 296p.
- Drewlow, L. W., P. D. Ascher and R. E. Widmer, 1973, Genetic studies of self-incompatibility in the garden chrysanthemum, Chrysanthemum morifolium Ramat, Theoret. Appl. Genet., 43:1-5.
- Leffel, R. C., 1963, Pseudo-self-compatibility and segregation of gametophytic selfincompatibility alleles in red clover, Trifolium pratense L., Crop Sci., 3:377-380.
- Riley, H. P., 1933, Self-sterility in Nemesia, Science, 77:326-327.
- Townsend, C. E., 1966, Self-compatibility response to temperature and the inheritance of the response in tetraploid alsike clover, Trifolium hybridum L., Crop Sci., 6:409-411.
- Whiting, G. F. and P. D. Ascher, 1974, Self-incompatibility in several Rhododendron species, Unpublished manuscript.