The Rhododendrons of North Korea
Jir Dostlek
Institute of Botany
Academy of Sciences of the Czech Republic
Pruhonice near Praha, Czech Republic
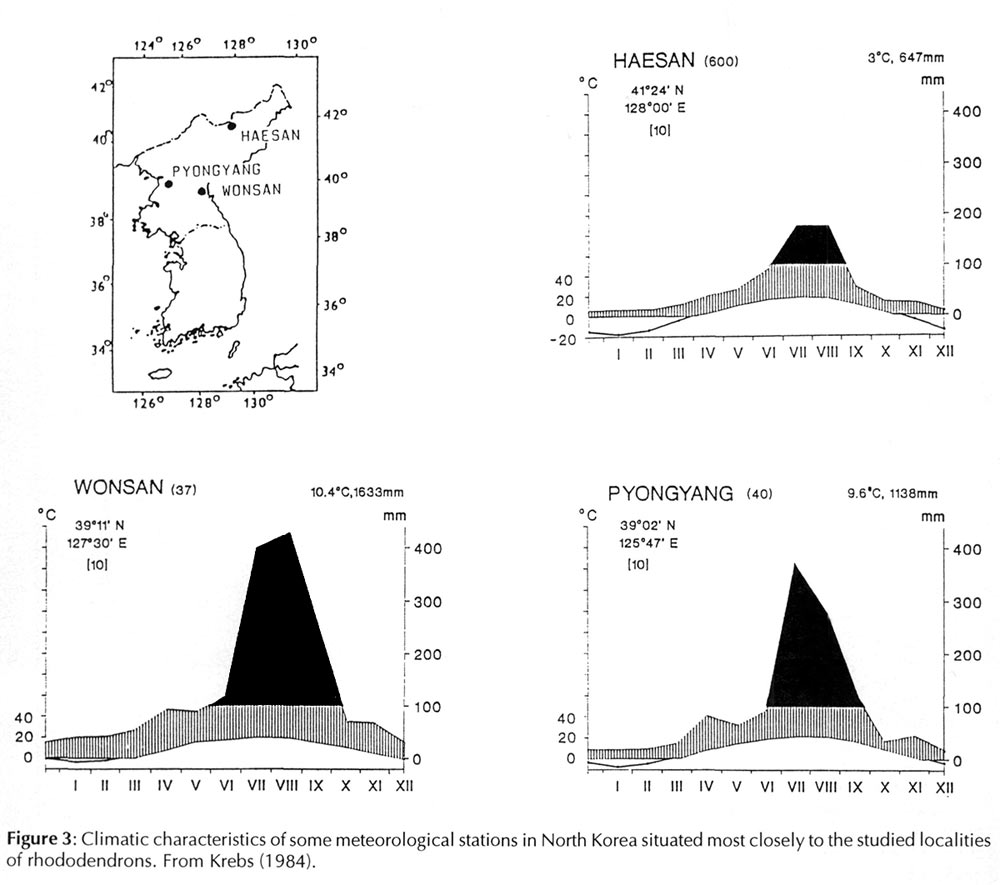
The northern border of the Korean peninsula lies on the same parallel as Rome, and its southern edge has the same latitude as Bagdad. From that one might assume that the North Korean climate will be quite warm. Latitude is quite misleading. In winter, predominantly northwestern, dry winds bring frigid weather from the Asian continent and can drop temperatures to -40°C. Beside that, the cold sea currents from the northeast contribute to the formation of the Korean climate in a similar but opposite manner than the warm Gulf Stream influences European climate. Korea is therefore very cold in winter and warm in summer. Of course, the climate also changes from the North to the South, which is reflected in native plant communities. Coniferous trees predominate in the northern part of the Korean peninsula, deciduous trees in the middle, and subtropical evergreen forests and cultivated citrus trees in the south.
The flora of North Korea is very rich, including many pre-Ice Age relicts because the glaciers stopped near the Chinese border. There occur many trees and shrubs which we cultivate in our gardens and parks, e.g., species of
Magnolia
,
Syringa
,
Rhododendron
,
Weigela
,
Philadelphus
,
Deutzia
and
Thuja
. This rich flora is exploited by North Korean gardening specialists, who use these species in landscaping around numerous and frequently huge monuments. They often dig up nearby plants, e.g., grown-up rhododendrons, and seedlings are available without lengthy cultivation.
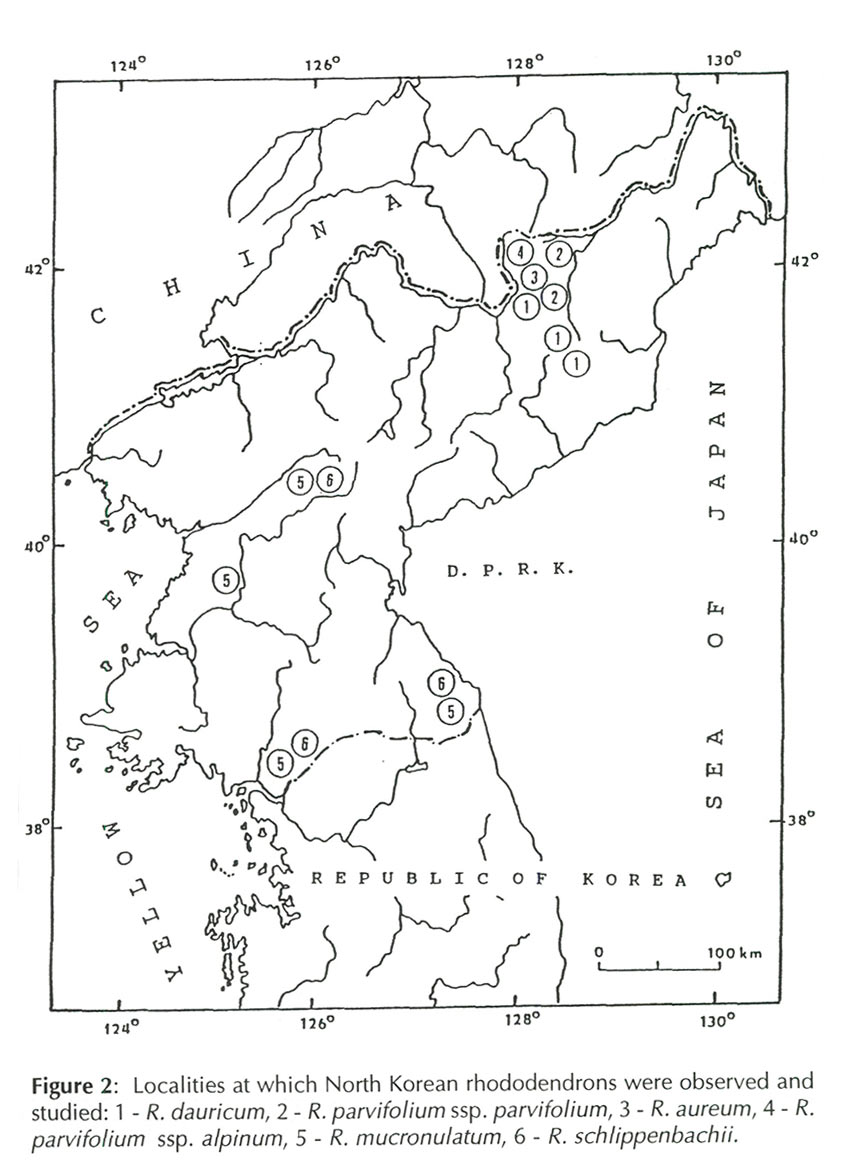
According to the Flora Coreana (Anonymous 1979), 13 species of the genus
Rhododendron
can be found on the Korean peninsula, namely
R. aureum
Georgi,
R. confertissimum
Nakai,
R. dauricum
L.,
R. faurie
Franch. (see Notes below),
R. micranthum
Turcz.,
R. mucronulatum
Turcz.,
R. parvifolium
Adams,
R. schlippenbachii
Maxim.,
R. tschonoskii
Maxim.,
R. weyrichii
Maxim.,
R. yedoense
Maxim. ex Regel and
R. redowskianum
Maxim.
Rhododendron confertissimum
Nakai is synonymous with
R. parvifolium
subsp.
alpinum
(Glehn) Dostlek et al. (2). During our expedition (May & June 1984) we found six species, including,
R. aureum
,
R. dauricum
,
R. mucronulatum
,
R. parvifolium
,
R. schlippenbachii
, and
R. yedoense
.
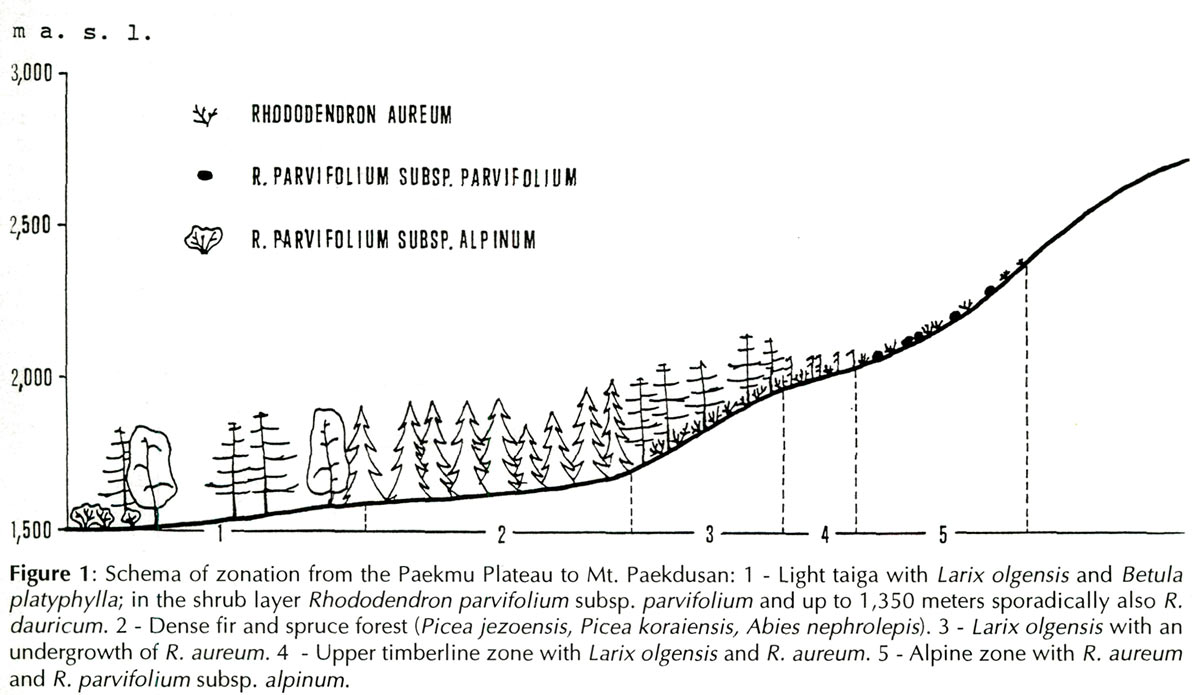
On the way from the capital of North Korea, Pyongyang, toward a mountain range on the Sino-Korean border and its highest mountain - volcano Paekdusan, we first saw the purple-pink flowers of
R. dauricum
on the mountain sides during our ascent of the mountains near Haesan town. With some interruptions they continued to be seen as far as the Paekmu Plateau, i.e., from about 800 to 1,350 m above sea level. This was on May 31, and it was as late as the first days of June when
R. dauricum
finished blossoming in this cold region. In Central European gardens this species blossoms, depending on the weather, in March or sometimes in April and exceptionally as early as in fall. In Korea it reaches a height of more than two meters. On some plants there occur non-deciduous leaves but only a few. Their total number is somewhat less than
R. dauricum
subsp.
ledebourii
(Pojark.) Alexandrowa et Schmidt (syn.
R. dauricum
var.
sempervirens
Sims) or even with
R. dauricum
subsp.
sichotense
(Pojark.) Alexandrowa et Schmidt (8), indicating that the North Korean population is
R. dauricum
subsp.
dauricum
.

|
|
R. dauricum
flowering on the foothills of the
Changbaishan Mts. at an elevation of ca. 800 meters. Photo by Jir Dostlek |
Rhododendron dauricum
occurs scattered in the undergrowth of forests dominated by
Abies nephrolepis
or
Larix olgensis
, accompanied by the following deciduous trees and shrubs at 800 m above sea level:
Betula platyphylla
,
Acer ukurunduense
,
Acer ginnala
,
A. tegmentosum
,
Acanthopanax sessiliflorum
,
Sorbus amurensis
,
Lonicera caerulea
,
Corylus heterophylla
,
Tilia amurensis
,
Ulmus laciniata
,
Schisandra chinensis
, and
Sorbaria sorbifolia
var.
stellipila
. At about 900 m elevation
R. dauricum
forms an almost continuous stand in the undergrowth of
Larix olgensis
forests. We observed the upper boundary of its occurrence on the Paekmu Plateau at about 1,350 m elevation, where it grows scattered in a larch taiga similar to plant communities including
R. parvifolium
Adams subsp.
parvifolium
(see Notes below). However,
Betula platyphylla
prevails over the larch in the tree layer and
Ledum palustre
var.
maximum
and var.
augustrum
occur in increased numbers in the shrub layer (see Notes below).
The soil in the root zone of
R. dauricum
at 800 m elevation is strongly acid, highly skeletal (89.8%) (see Notes below), and nutrient poor with a high proportion of organic matter (29.4%). At the altitude of 1,350 m elevation, on the basalt Paekmu Plateau, the soil is quite similar; only the content of organic matter varies according to the site between 3.6 and 28.4%.
|
On the Paekmu Plateau, at elevations starting from about 1,100 m R. parvifolium subsp. parvifolium grows beside R. dauricum . There it is much more common and extends somewhat higher than the previous species, namely up to 1,500 m. It is 70 to 130 cm tall, erect, with non-deciduous leaves only 12 to 27 mm long; individual flowers are about 25 mm in diameter and are red-purple. Dwarf and prostrate plants do not occur there. From June 3 to 5, shrubs were in full bloom. This subspecies is widespread in the larch taiga, especially in its glades, where it grows together with shrubby Betula paishanensis , Lonicera caerulea and Vaccinium . The trees Betula platyphylla and Prunus padus encroach into the glades. Rhododendron parvifolium subsp. parvifolium is often dominant in waterlogged habitats and small swamps, with willows and among the herbs, especially sedges. These stands are beautiful in bloom and inspiring for eventual use in landscaping.

|
|
R. parvifolium
subsp.
alpinum
on Mt. Paekdusan.
Photo by Jir Dostlek |
On Mt. Paekdusan, above the timberline, in the alpine zone at 2,050 to 2,400 m elevation, we found a very nice dwarf R. parvifolium subsp. alpinum (Glehn) Dostlek et al. It is only 2 to 5 cm tall and prostrate and has leaves only 8 to 18 mm long and individual flowers 20 mm in diameter. It does not grow larger even at its lowest and most protected sites. It is spatially remote from the erect and much larger R. parvifolium on the Paekmu Plateau, separated by a barrier of evergreen forest, composed of firs and spruces ( Picea iezoensis , P. koraiensis , Abies nephrolepis ). On June 3 to 5, when we visited the locality, subsp. alpinum was in full bloom. Only at the highest elevations ca. 2,400 m elevation were there certain noticeable differences among individual plants: some flowered, others had not. Above the timberline, R. parvifolium subsp. alpinum together with R. aureum form dwarf-shrubby communities with several willow species and also with Vaccinium vitis-idaea , Vaccinium uliginosum agg., Phyllodoce caerulea and Juniperus sibirica . In higher altitudes, from 2,200 m, the stands open.

|
|
Detail of an inflorescence of
R. parvifolium
subsp.
parvifolium
on the Paekmu Plateau, about 1,400 meters. Photo by Jir Dostlek |
Rhododendron parvifolium subsp. parvifolium grows on the Paekmu Plateau on the same soils as does R. dauricum from the same localities (see above). The alpine subspecies R. parvifolium subsp. alpinum grows together with R. aureum on soils composed largely of pumice gravel saturated to a various extent with organic matter from decomposed plants. The soils are acid to strongly acid, poor in nutrients and have a high content (40.9 to 72.7%) of skelet and of organic matter from 11.9 to 31.9%.

|
|
R. parvifolium
subsp.
parvifolium
on the
Paekmu Plateau, about 1,400 meters. Photo by Jir Dostlek |
As mentioned earlier, R. parvifolium subsp. alpinum grows above the timberline together with R. aureum . But R. aureum encroaches below the timberline at an elevation of 200 m. It grows at elevations of 1,700 m up to almost the upper boundary of the alpine vegetation zone at 2,400 m. At the upper edge of this locality, in leeward wettish furrows, R. aureum is only 5 cm tall. At 1,700 m elevation the dense evergreen forest of spruces and firs ends, the stand opens and a larch, Larix olgensis var. koreana with a total coverage of 30-40% succeeds as the dominant tree. There R. aureum is abundant, growing up to 70 cm tall. Morphologically, the small plants from the upper part of the site and the taller ones from the lower part are identical; here we were dealing only with the influence of more favorable site conditions.

|
|
R. aureum
as a dominant species in the undergrowth
of Larix olgensis on Mt. Paekdusan, just below the timberline, about 1,800 meters. Photo by Jir Dostlek |
Below the timberline,
R. aureum
forms a distinct association with larch (see Dostlek at al., 1988). In the herb and low shrub layer, we found
Phyllodoce caerulea
,
Diphasiastrum complanatum
,
Juniperus sibirica
,
Linnaea borealis
,
Vaccinium uliginosum
,
Ledum augustissimum
(see Notes below),
Lonicera caerulea
,
Clintonia udensis
and
Viola sachalinensis
.
The soils there are strongly acidic and nutrient poor with relatively high contents of skelet and organic matter, mostly residues from decaying needle-leaves and herbaceous plants. Above the timberline, at an elevation of about 2,000 m,
R. aureum
forms compact dwarf-shrub communities. At its highest elevation (about 2,400 m), in places with a favorable relief, such as small depressions, leeward sites or trenches carved by erosion in pumice gravel, it forms uniform or monotypic stands or occasionally associates with dwarf shrubs of
Phyllodoce caerulea
.

|
|
R. aureum
in the alpine zone of Mt. Paekdusan, about 2,400 meters.
Photo by Jir Dostlek |
On June 3 to 5,
R. aureum
was in full bloom. The light-yellow flowering display both in the alpine zone and under larches, makes an unforgettable impression.
Rhododendron aureum
is very closely related to the Caucasian species,
R. caucasicum
Pall. Mt. Paekdusan volcano, whose slopes and foothills
R. aureum
inhabits, is 2,744 m high and was last active in 1702. According to the remnants of trees from that time, which here and there emerge from volcanic ash, the upper timberline used to be higher, and today's vegetation has not yet completely recovered to those high elevations.
We familiarized ourselves with
R. mucronulatum
and
R. schlippenbachii
, mainly in the Myohyangsan and Kumgangsan mountains. The name "Myohyangsan" means "sweet-smelling mountains" in English, perhaps because of the smell of the flowers which grow there. The Kumgangsan Mts. (diamond mountains) form one of the most beautiful natural Korean landscapes. Both of the above-mentioned species are widespread and often grow together. We occasionally saw them as well at other locations. They grow at elevations between 50 and 1,400 m (
R. mucronulatum
up to 1,600 m) and can grow up to 3 m tall. Beside
R. mucronulatum
var.
mucronulatum
, which has sparse tiny scales on its leaves, we found, mainly in the Kumgansan Mts., the variety
ciliatum
Nakai, which in addition to the tiny scales also has sparse simple hairs on its leaves and cilia on the margins of its leaf blades. There also exist transitional forms. Both species had already passed their blooming periods when we saw them for the first time in June. On June 16, only in the upper part of Mt. Wonmanbong in the Myohyangsan Mts., at elevations of about 1,000 to 1,200 m, were some shrubs of
R. schlippenbachii
just finishing their bloom.
Rhododendron schlippenbachii
was found in a forest of
Quercus mongolica
, a very interesting dominant and compact shrub layer.
The
R. mucronulatum
and
R. schlippenbachii
occur most frequently in thin, light forests. In the lower reaches of the Myohyangsan Mts. (300 to 500 m) they occur scattered through the pine forests dominated by
Pinus densiflora
. From an elevation of 500 to 600 m, a "blueberry" with red berries,
Vaccinium koreanum
, joins the understory and in this community both rhododendrons are more common; at higher altitudes
R. schlippenbachii
is most common in stands with
Quercus mongolica
. In the Kumgangsan Mts., both species grow most often in forest communities with this particular oak.
Rhododendron mucronulatum
is more common than is
R. schlippenbachii
in unshaded habitats, e.g., in rock cracks on dry sunny mountain summits. There it can display a very stunted form.
The soils, which developed on igneous bedrocks or their metamorphisms, do not substantially differ from the situation described earlier in connection with other species. Only in the Myohyangsan Mts. we found soils that were especially deficient in magnesium.
| Table 1 : Chemical and physical composition of the soil samples (contents in mg per kg dry mass [p.p.m.], except organic matter and gravel as percentages of dry mass). Legend: D - R. dauricum, Pp - R. parvifolium subsp. parvifolium, Pa - R. parvifolium subsp. alpinum , A - R. aureum , M - R. mucronulatum , S - R. schlippenbachii . From Dostlek et al. (1988). | ||||||||||||||
| Loc. no. | Species | pH | Na + | K + | Mg 2+ | Ca 2+ | N-NH 4 2 | N-NO 3- | N-total | P-PO 4 3- |
Org. matter
(%) |
gravel
>2 mm (%) |
||
| In H 2 O | In KCI | |||||||||||||
| 1 | D | 4.60 | 3.55 | 120 | 26 | 380 | 1,000 | 55 | 4.0 | 7,700 | 10 | 29.41 | 89.8 | |
| 2 | Pp | 5.20 | 4.55 | 70 | 120 | 80 | 700 | 60 | 4.0 | 3,080 | 8 | 28.38 | 81.5 | |
| 3 | D | Pp | 5.60 | 4.90 | 65 | 80 | 48 | 300 | 23 | 2.6 | 2,100 | 6 | 13.93 | 89.7 |
| 4 | Pp | 4.95 | 4.50 | 85 | 100 | 60 | 300 | 35 | 3.0 | 1,680 | 10 | 3.61 | 75.7 | |
| 5 | A | 4.30 | 3.60 | 95 | 126 | 220 | 800 | 55 | 7.4 | 4,760 | 6 | 29.41 | 70.3 | |
| 6 | A | Pa | 5.55 | 4.95 | 90 | 170 | 240 | 1,400 | 45 | 3.6 | 5,040 | 11 | 11.87 | 40.9 |
| 7 | A | Pa | 5.05 | 4.70 | 125 | 126 | 180 | 1,700 | 48 | 5.0 | 4,060 | 7 | 27.86 | 53.2 |
| 8 | A | Pa | 5.20 | 4.50 | 130 | 270 | 360 | 1,400 | 75 | 3.6 | 6,860 | 11 | 30.96 | 72.7 |
| 9 | M | S | 5.15 | 4.65 | 70 | 120 | <1 | 200 | 40 | 2.0 | 2,380 | 4 | 27.35 | 72.4 |
| 10 | S | 4.70 | 4.55 | 60 | 170 | <1 | 160 | 40 | 2.0 | 2,660 | 5 | 40.25 | 55.2 | |
| 12 | M | S | 4.45 | 3.95 | 75 | 80 | 52 | 200 | 40 | 4.4 | 3,080 | 8 | 18.58 | 88.9 |
|
Localities:
(1) Province of Ryanggang, Naegok, NW slope above the local spa resort, altitude 800 m, 4133', N and 128°25', E; June 1, 1984. (2) Prov. of Ryanggang, NW edge of Peakmu Plateau, altitude 1,500 m 41°57', N and 12818', E; June 3, 1984. (3) Prov. of Ryanggang, N part of the Peakmu Plateau , near the Mubong Hill, altitude 1,350 m 41°57', N and 128°32', E; June 5,1984. (4) Prov. of Ryanggang, NE part of the Peakmu Plateau, near Taehongdan, altitude 1,100 m, 4157', and 12840', E; June 5 1984. (5) Prov. of Ryanggang, the Changbainshan Mts., E slope of the Mt. Mubudong, altitude 1,850 m,4157', N and 1 28°11', E; June 3 and 5, 1984. (6) Prov. of Ryanggang the Changbaishan Mts., SE foot of the Mt. Peakdusan (above timberline), altitude 2,100 m, 41°58', N and 12808', E; June 3 and 5 1984. (7) Prov. of Ryanggang, the Changbaishan Mts., SE slope of the Mt. Paekdusan, altitude 2,200 m, 41°59', N and 128°06', E; June 3,1984. (8) Prov. of Ryanggang, the Changbaishan Mts., SE slope of the Mt. Paekdusan, altitude 2,400 m, 4159', N and 1 28°08', E; June 3, 1984. (9) Prov. of North Pyongan, the Myohyangsan Mts., S slope below the Sangwonam Monastery Complex, altitude 520 m, 40°01', N and 126°18', E; June 14, 1984. (10) Prov. of North Pyongan, the Myohyangsan Mts., S slope E of the Kusungpokpo Waterfall, altitude 600 m, 4001', N and 12619', E; June 15, 1984. (11) Prov. of North Pyongan, the Myohangsan Mts., NNW slope above the Myohyangsan Hotel, altitude 300 m, 40°01', N and 12618', E; June 18, 1984. (12) Prov. of Kangwon, the Kumgangsan Mts., (the Diamond Mts.), SE slope of the Mt. Chonsondae, altitude 870 m, 38°45', N and 128°05', E; June 30, 1984. |
||||||||||||||
Rhododendron yedoense var. yedoense is cultivated in Korea, which by the end of May had practically finished blooming in gardens. In the field we did not come across R. yedoense var. poukhanense (Lev.) Nakai, which is the wild type from which var. yedoense arose.
|
|
Rhododendrons in regions distant from each other have long been used in folk medicine, often for the same purposes. In Korea the flowers of R. mucronulatum are used, and the leaves of R. fauriei are recommended as diuretic and diaphoretic preparations and as a remedy against high blood pressure.
Notes
Hultn (1958, 1968) considered
R. parvifolium
as synonymous with
R. lapponicum
(L.) Wahlenb., and M.N. Philipson and W.N. Philipson (1975) concurred. This opinion has its predecessors, because as early as 1872 Herder joined
R. parvifolium
to
R. lapponicum
(
R. lapponicum
var.
parvifolium
). The similarity of
R. lapponicum
particularly with
R. parvifolium
subsp.
alpinum
is striking. If we accept the view that we are not dealing with convergence but with identity of both species and if we consider them according to the rule of priority as
R. lapponicum
(L) Wahlenb., then we should respect variability at the infraspecific level. Because, in North Korea, the difference between the two subspecies of
R. parvifolium
Adams is conspicuous and clear-cut. There occurred there a pronounced differentiation of ecotypes in separate localities. Our first impression in the field was such that one could understand why Nakai considered subsp.
alpinum
to be an independent species (
R. confertissimum
Nakai) and that this view was accepted even by Korean botanists. Transitional forms do not occur there. Beside the different intensity of growth and the upright vs. horizontal orientation of the stems, which are not so significant from the taxonomic viewpoint, the subspecies have markedly different ecology. Subspecies
parvifolium
grows in forests and forest glades and has a dominant position on waterlogged soils and small peat swamps, whereas subspecies
alpinum
grows above the timberline on pumice gravel. Growth characteristics and stem orientation of both subspecies of
R. parvifolium
Adams were verified under cultivation, and it was confirmed that they are genetically encoded. To reconsider the problem and revise current views we should evaluate populational variability throughout the whole range of distribution and eventually determine whether or to what extent the "ecobiomorphs" mentioned by Mazurenko (1980) are based on heredity.
Nomenclature of species generally follows Flora Coreana (Anonymous 1979) In this article the problems of the genus
Ledum
reclassification into the genus
Rhododendron
and the problems of synonyms of
R. fauriei
with
R. brachycarpum
are not solved.
"Skelet" (inorganic material in the soil, e.g., stones, gravel).
Acknowledgements
My thanks are due to Dr. Mark P. Widrlechner for his help with my English, to Frank Doleshy for his comments on the manuscript and to Sonja Nelson for her help in preparation of the manuscript for press.
Literature Cited
1. Anonymous. Flora Coreana, appendix. Pyongyang; 1979.
2. Dostlek, J. Sr.; Dostlek, J. Jr.; Mucina, L.; Hoang, Ho-Dzun. On taxonomy, phytosociology and ecology of some Korean
Rhododendron
species. Flora 181: 29-44; 1988.
3. Hultn, E. The amphi-atlantic plants and their phytogeographical connections. Kungl. Svenska Vetenskapsakad. Handl., Stockholm, ser. 4., 7(1): 200; 1958.
4. Hultn, E. Comments on the flora of Alaska and Yukon. Arkiv. Bot., Stockholm, 7(1): 93; 1968.
5. Krebs, J. M. et al. World Weather Records 1961-1970, Asia, vol 4., p. 92-99. Asheville, N.C., USA; 1984.
6. Mazurenko, M. T. Rododendrony dainego vostoka. Moskow; 1980.
7. Philipson, M. N.; Philipson, W. R. A revision of Rhododendron section Lapponicum. Not. Roy. Bot. Gard. Edinburgh 34: 1-71; 1975.
8. Schmidt, P.; Alexandrowa, M. S. Die wildwachsenden Rhododendron-Arten der UdSSR. - Beitr. Gehzkunde 1981: 55-76; 1982.