Rhododendron 'Sri Chinmoy'
John L. Rouse and Leslie A. Bursill
School of Physics, The University of Melbourne
Parkville, Victoria Australia
Introduction
Rhododendron loranthiflorum
Sleumer is a tropical, epiphytic,
Vireya
species with white perfumed flowers found on the island of Bougainville in the Solomon Islands at an altitude of about 1000m. In Melbourne, a number of specimens have been growing out of doors in the garden of John Rouse for the last 25 years. One particular clone is outstanding in that it is easily cultivated, grows as a dense shrub and is highly floriferous with some flowers to be found at most times of the year.
Here we give a brief introduction to the species and its natural habitat and describe our outstanding specimen that has been named after the New York guru Sri Chinmoy to honour his international endeavours to attain world peace.
The tropical vireyas are naturally distributed in the equatorial region bounded by the Philippines in the north, Sumatra and Peninsular Malaysia in the west, North Queensland in the south and the Solomon Islands in the east, with a concentration of species diversity found on the islands of New Guinea and Borneo. These two regions of species concentration are separated by Wallacea (Wallace, 1869), the biologically discontinuous region which contains the line of join between Eurasia and the Australian plate when they collided some 10 million years ago and brought together their own groups of animals and plants. The tropical vireyas, which probably arose from a wandering Himalayan ancestor, have developed over the past three or four million years with considerable isolation between the species east and west of Wallacea. Mostly, the tropical vireyas are found in the mountains at altitudes of 1000 to 4000m above sea level, so while day and night times are equal the year round and relative humidity is high, the temperature is lower than that at tropical sea level.
Rhododendron loranthiflorum
is a scaly species in subgenus
Rhododendron
, section
Vireya
, subsection
Solenovireya
(Sleumer, 1966). This subsection contains over 30 species spread out from Sumatra and Peninsular Malaysia to Bougainville but excluding Java and Australia. Their characteristics include well spread out flowers with a tubular corolla and a relatively long, straight or slightly curved tube. This collection of species appears to lack cohesion in that their diverse properties suggest that they should be divided into two or more taxa (Stevens, 1985; Williams and Rouse, 1997).

|
|
Figure 1. A plant of
R. loranthiflorum
'Sri Chinmoy' photographed at the
Annual Show of the Australian Rhododendron Society, Olinda Gardens, when it was awarded the Certificate of Merit in November 1974. Photo by John L. Rouse |
R. loranthiflorum
'Sri Chinmoy'
Seed of
Rhododendron loranthiflorum
was collected by Lyn Craven in 1964 near Lake Loloru on Bougainville at about 1000m with the collection number craven (SCHODDE) 158 and distributed to members of the Australian Rhododendron Society. In December 1970 Lyn gave John Rouse two cuttings of
R. loranthiflorum
, one of which rooted and developed rapidly, so that in November 1974 as a container grown shrub in full flower, he entered it in the competitive display at the Annual Show of the Australian Rhododendron Society as "plant in flower of any species or hybrid of the Malesian group" (Figure 1). There it obtained first prize and was awarded the Certificate of Merit by the judges. In 1997, when this rhododendron had reached a height of nearly 8 ft.(2.4m) and regularly produced a magnificent display when covered in flowers, it was registered with the name
R. loranthiflorum
'Sri Chinmoy'. While the inflorescence of other forms of
R. loranthiflorum
characteristically have 4 to 6 flowers, this form typically displays inflorescences containing 8 or 9 flowers (Figure 2).

|
|
Figure 2. An inflorescence of
R. loranthiflorum
'Sri Chinmoy' showing
8 flowers with 7 leaves in the pseudowhorl below. The photograph was taken with a 6 x 6 cm format Rolleiflex SL66 camera whose lens could be angled up and down with respect to the film plane. The angle used was 4 down and this was of great assistance in focusing a very open 3-dimensional truss. A pair of flash lamps provided the illumination. Photo by John L. Rouse |
A number of plants of
Rhododendron loranthiflorum
, raised from seed received from Tom Lelliot in November 1974, are also growing in the garden, but although their flowers are similar to those of our named specimen, the plants are more difficult to maintain in cultivation; they grow more slowly and are less floriferous.
Rhododendron luraluense
Sleumer, in subsection
Euvireya
, is also found on Bougainville where it grows as a small epiphytic shrub. Our garden specimens have reached a height of about 1 m and their flowers are white and attractive (Figure 3). However, this species seems difficult to keep alive and the surviving plants are grafted onto F1 vireya hybrids with terrestrial parents.

|
|
Figure 3. An inflorescence of
R. luraluense
, whose white flowers have a
broader and shorter corolla tube and wider lobes than R. loranthiflorum . Photo by John L. Rouse |
Pollinators
To be certain how rhododendrons in the wild are pollinated requires observation on visitors to the flowers, whether the visitors transfer pollen between flowers and whether viable seeds of the species are produced. For epiphytes high up in the trees, such observations are not a trivial exercise. Further, some species with red flowers, such as
Rhododendron lochiae
which inhabits isolated mountain tops in north Queensland, have characteristics in the UltraViolet-A which suggest that although they are primarily bird pollinated they have a possible back-up insect pollinator. Such speculation can only be confirmed by direct observation. However, a guide to possible pollinators can be obtained from characteristics of the flowers and inflorescence. In particular, the reflection characteristics of the corolla in the UltraViolet-A (wavelength 320 to 400 nm) can be significant, as in general insects can see in the UV-A but are red blind, while birds and bats can see in the red but not in the UV-A.

|

|
|
|
Figure 4(a). The face of the corolla of
R. loranthiflorum
'Sri Chinmoy' photographed with UV-A illuminators. Photo by John L. Rouse |
Figure 4(b). The same face photographed with two Spectroline E14F
UV-A illuminators mounted close to the camera lens, a Kodak Wratten filter type 18A over the lens which transmits UV-A but not visible light, Kodak Tri-X-Pan film and exposure of 8 seconds at f16. Film processing and printing were standardised and one frame on each film was used to photograph in visible standardised light a chart composed of white card, grey neutral density card and the black cartridge paper used as the back-drop when photographing the flower with ultra-violet illumination. The corolla is only a partial reflector in the UV-A. Photo by John L. Rouse |
Figures 4(a) and (b) show the face of the corolla of Rhododendron loranthiflorum 'Sri Chinmoy' as photographed in visible light and UV- A respectively. The dark centre in 4(a) is because the light did not reach the inside of the long thin corolla tube. In contrast to visible light, Figure 4(b) shows that the UV-A radiation was partially absorbed by the corolla face except close to the centre where the absorption was high. Visible "bee guides" to attract insect pollinators can often be seen on rhododendron corollas. However, while vireyas show no such visible guide, they frequently display in the UV-A a central absorption guide as we see here. It is probable that taking into account the white corolla with small lobes, its long thin tube and UV guide, the aroma and upright presentation of the flowers, that its main pollinators on Bougainville are Sphingid moths (Stevens, 1976).
Sexual Compatibility
When self-pollinated,
Rhododendron loranthiflorum
'Sri Chinmoy' sets viable seed, which may produce some offspring very similar to their parent. In the mid-1970s, selfed seed was sent to vireya enthusiasts in the USA, Europe, Japan and New Zealand as well as within Australia and cuttings were sent to the USA. It is our view that even with similar appearance, selfed seedlings should be called
"R. loranthiflorum
'Sri Chinmoy' selfed" to distinguish them from their parent. In the artificial environment of our Melbourne garden, there are apparently no effective pollinators for
R. loranthiflorum
, as this species does not set seed unless hand-pollinated.
The style length is about 31mm and cross-pollinations can be made within tropical vireyas that result in F1 hybrids provided the style length of the other parent is not too dissimilar (Williams and Rouse, 1988). The following successful crosses were made in the 1970s and selected hybrids are growing in the garden:
R. loranthiflorum
'Sri Chinmoy' x
R. konori
(Irian Jaya form)
R. macgregoriae
x
R. loranthiflorum
'Sri Chinmoy' (Figure 5)
R. loranthiflorum
'Sri Chinmoy' x
R. laetum
(Figure 6). These hybrids are robust, vigorous plants which thrive in full sunlight for much of the day and grow to a height of up to 2 to 3m. As F1 hybrids they show characteristics of both parents and are closer to the "average vireya" than either parent so that the advantages of hybridisation will only become apparent when selected forms of the F2s are developed.
Cross-pollinations between tropical vireyas and rhododendrons from other taxa seldom result in viable seed, and on the few occasions when a hybrid is produced it is completely sterile and of little horticultural value (Rouse et al., 1993).

|

|
|
|
Figure 5. An inflorescence of the hybrid
( R. macgregoriae x R. loranthiflorum 'Sri Chinmoy') Photo by John L. Rouse |
Figure 6. An inflorescence of the hybrid
( R. loranthiflorum 'Sri Chinmoy' x R. laetum ). Photo by John L. Rouse |
Graft Compatibility
The species in subsection
Soleno-vireya
graft compatibly on to hybrid
Vireya
stock, but their behaviour when grafted on to
Rhododendron
'Fragrantissimum' ([
R. edgeworthii
x
R. formosum
], both in section
Rhododendron
and from the Himalayan region) is variable. In general, graft compatibility of
Vireya
species on
R.
'Fragrantissimum' depends on whether they come from regions to east or west of Wallacea; those from the west are compatible, those from the east are not (Rouse and Williams, 1997), but the variability arises because species in subsection
Solenovireya
are less likely to conform to this pattern than species from other subsections. Since
R. loranthiflorum
'Sri Chinmoy' grows well on its own roots at least in Melbourne, it is rather academic how it performs when grafted. However it is incompatible long-term on
R.
'Fragrantissimum'.
Symmetries
Symmetry in plants, called phyllotaxis, has been investigated for centuries usually by botanists or mathematicians and frequently the Fibonacci sequence of integers <0, 1, 1, 2, 3, 5, 8, 13, 21, ...> appears in the discussion. Physicists are also interested in symmetry particularly the symmetries in solid inanimate matter. However, both the authors are physicists and we have expanded our interest in symmetry to include the study of symmetries in mature plants and in particular a survey of the
Rhododendron
species (Bursill and Rouse, 1998).
Most of the tropical vireyas have a few leaves (3 to 20) arranged in a pseudowhorl (some exceptions are
R. ericoides
,
R. quadrasianum
and
R. womersleyi
) and
Rhododendron loranthiflorum
has a pseudowhorl with about 5 leaves. The term "pseudowhorl" means that while the leaves are close to the same height up the stem they do in fact have a definite order from the lowest to the highest so they are not whorled. However, fluctuations in height possibly due to the closeness of their petioles on the stem, may result in some leaves being out of order. There are also other organs arising from the mature stem the most obvious being the flower pedicels at the top. In between there are the flower bud scales, which have usually abscised by the time the flowers open. Below the pseudowhorl of leaves there is a longer length of stem that is without leaves, though on close inspection it may show scars where there were a few small leaflets when the stem first developed.
Phyllotaxis is the investigation of the arrangement of these organs (leaflets-leaves-bud scales-flowers) on the stem, the production of an ideal mathematical model to represent a particular stem and the comparison between the real stem and this model. Thus, the first requirement is to determine the location of each of these particular organs on the stem by measuring the local stem diameter Dn mm, their height up the stem Hn mm and their angle about the stem axis dn degrees where n = 0,1,2, ...N refers to the nth particular organ and N + 1 is the total number of each of these four groups of organs.
Figure 7 shows two photographs of the F1 hybrid
Rhododendron christianae
x
R. jasminiflorum
used here as a replacement for
R. loranthiflorum
'Sri Chinmoy' which had recently flowered, so that no suitable floral stems were available because of the rapid decay of the peduncle. The leaves and flowers have been removed from the hybrid stems so that only their scars remain. The hybrid's two parents have similarities to
R. loranthiflorum
and the hybrid stem is very similar to that of
R. loranthiflorum
though it is slightly larger. Indeed, this hybrid acts as a guide to those tropical vireyas that have a pseudowhorl of leaves, as could be expected, since most of the characteristics which distinguish
Vireya
species from each other have been removed; i.e., for the flowers - shape, colour, aroma, size, pedicel length and presentation, and for the leaves - shape, venation, size and petiole length. What is left includes stem diameter which suggests size, the scale type which gives a guide to the subsection, colour and, our interest, the number and arrangement of the leaflets, leaves, bud scales and flowers on the stem or peduncle. Figure 7 includes two stems from the hybrid as the top leaflet scar is only seen in one photograph while the other gives clearer details of pedicel and flower bud scale scars.
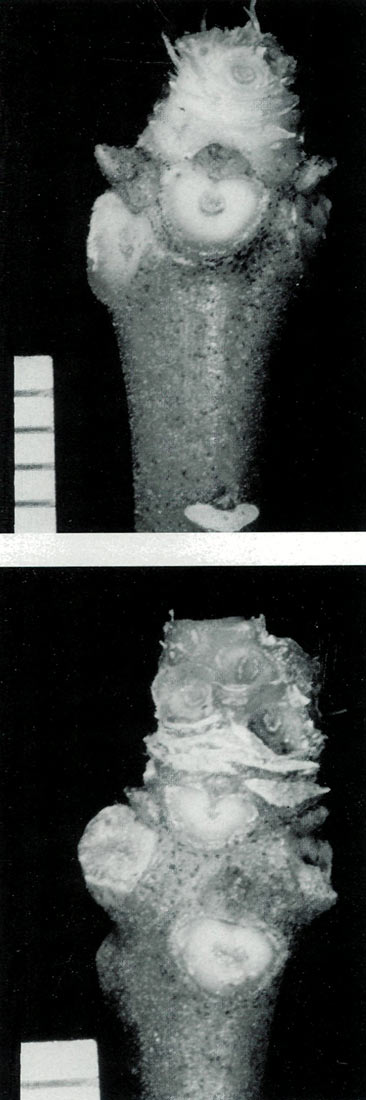
|
|
Figure 7. Two mature stems of the vireya
hybrid ( R. christianae x R. jasminiflorum ) with leaves and flowers removed as a replacement for R. loranthiflorum 'Sri Chinmoy'. Figure 7(a) shows a scar at the bottom, left behind when the top leaflet abscised, and Figure 7(b) shows the scars left by the bud scales. Both figures show the scars seen when the pseudowhorl of leaves was removed and at the top, the scars on the peduncle where the flower pedicels were attached. The scale intervals represent 1 mm. Photo by John L. Rouse |
Rhododendron loranthiflorum 'Sri Chinmoy' has a normal mature stem with 30 to 50mm bare except for 0 to 10 leaflet scars, 3 to 6mm of stem usually with 5 leaves in the pseudowhorl, 1mm of bud scales and 2 to 3mm of flower pedicels at the top arising from a dome shaped peduncle. When the plant is given excessive fertiliser, the leaflet region is stretched to 100 to 150mm, the number of leaves increases to 10 to 15 and the flower bud is replaced with a growth bud.

|
|
Figure 8. A photograph from above of a mature stem of
R. loranthiflorum 'Sri Chinmoy' which has flowered. The leaves are numbered with mapping pins with left pin = 0 the lowest leaf, upper right pin = 1, bottom pin = 2, upper pin = 3, lower right pin = 4 and the 6th small leaf without a pin. Photo by John L. Rouse |
Figure 8 shows a stem of
Rhododendron loranthiflorum
'Sri Chinmoy' photographed from above after flowering and with mapping pins through the leaves indicating the order of the leaves up the stem (their n value) with left pin 0, upper right pin 1, bottom pin 2, top pin 3 and lower right pin 4. There is a sixth leaf which is too small for a pin representing 5. Frequently as we see here, the lowest or highest leaves are small compared to those in the middle of the pseudowhorl. This photograph can be used to measure the angle between a leaf and the next one up the stem. In this example, it is about 135° (= 3/8 x 360° where 3 and 8 are from the Fibonacci sequence above with one number [5] in between) clockwise so that the leaves are arranged as a left-handed helix (compare with a corkscrew) on the stem and it takes 8 leaves to go just 3 times round the stem. Similarly, if the flower pedicels are present in the photograph, they can be used to measure the angle between one flower and the next one up the peduncle. Observation shows that the leaflets, leaves, flower bud scales and flowers all have this helical arrangement on the stem and that almost always they all have the same handedness on a particular stem (Bursill & Rouse, 1998). Left- and right-handed helices are mirror images of each other so they and the leaves on the stem they represent are chiral. Chirality from DNA molecules up to complete organisms are an important characteristic of life on earth but the chirality of a
Vireya
stem is not of significance and when lateral branches are formed, the chirality may change, so a clone or well-branched plant contains both left- and right-handed stems. However, Figure 8 shows that the leaf petioles do not regularly leave the stem radially so that there is some uncertainty in the angle associated with a particular leaf although the overall mean angular difference is accurate
Currently the authors are investigating the phyllotaxis of the leaves on
Lilium
stems up to 2.5m tall and with hundreds of leaves. We then plan to investigate in more detail the phyllotaxis of a number of vireya species including their pseudowhorls of 5 to 10 leaves where the equivalent stem length is only a few mm so we can compare the two systems. This requires accurate measurements using the vireya scars on the stem. The chief problems to be overcome are:
(a). The flower bud scale scars are small compared to other scars and they are old since the scales abscised not long after the flowers opened. To observe and measure these scars they need to be viewed with at least x4 magnification.
(b). Once the pedicels have been removed, their scars rapidly deteriorate, possibly distort and are difficult to distinguish. It thus appears necessary to remove the pedicels one at a time and measure the scar before removing the next pedicel.
(c). The petiole scars from the pseudowhorl, the bud scale scars and pedicel scars cover most of the surface of the stem (such close neighbours may well influence the position of the scar). It does mean that there is no bare stem on which to measure the stem diameter except just below the pseudowhorl and perhaps over the bud scale scars. As well, there is a tendency for stems to have non-circular cross-section just below the pseudowhorl, in which case we take the mean of the largest and smallest axis. We use a parallel jawed micrometer to measure the stem diameter, taking care not to deform or damage the soft stem.
Initial trial measurements on a well developed, left-handed stem of
Rhododendron loranthiflorum
'Sri Chinmoy' grafted on to
R. retusum
x
R. quadrasianum
gave D = 3.5mm for 40mm of stem with no leaflet scars and for 2 leaves before the pseudowhorl and D = 4mm just before the pseudowhorl and over the flower bud scale scars. Within the pseudowhorl of 10 leaves, the average angle between one leaf and the next one up the stem was 134.6 degrees and the average height between one leaf and the next one up the stem was 1.5mm. These figures were obtained after rearranging 3 leaves, which appeared out of order. The similar results for the 11 bud scale scars were 135.8 degrees and 0.13mm after reordering 3 scars and noting that a scar had been missed. During the above measurements, the flower pedicel scars deteriorated so they were not measurable. These average values after reordering give a model for the helices representing the arrangement of the leaves and bud scales on the stem, a model which can be compared with the original data points and further developed in ways of interest to mathematicians and crystallographers. The above measurements of dn and Hn have inadequate accuracy as they were made using our phyllotactic bench designed for
Lilium
. We are currently making modifications to the bench so that it will be suitable for measuring both
Lilium
and
Rhododendron
.
Cultivation
It is sometimes said that the tropical vireyas are hardly "tropical" since although they are equatorial, the climate at altitudes of 2000 to 4000m is as regards temperature temperate. While the word tropical is usually added to indicate that those subsection
Pseudovireya
species from temperate regions more than 20° north of the equator are to be excluded, the word tropical is appropriate as regards relative humidity (90 to 100%), day length (12 hours) with high rainfall and epiphytic growth. At 1000m on Bougainville, the climate is thoroughly tropical so that it is unexpected that
Rhododendron loranthiflorum
'Sri Chinmoy' would be easy to cultivate in the open in Melbourne (38° south). In the last two decades our screen air temperature has been in the range 0.5 to 44°C and the annual rainfall 430mm to 1000mm. In summer, high temperatures are frequently accompanied by strong dry winds from the north with a relative humidity of less than 10%. Experience suggests that vireyas do not like negative air temperatures and that - 4 to -5°C is usually sufficient to produce death. As well, they do not tolerate a frost on their leaves. We do have frosts with grass temperatures as low as -6°C but in the last decade frosts have occurred less frequently. Since the frosts only occur in the open, the vireyas are grown in frost-free areas where there is shelter from trees or buildings.
High temperatures above 38°C can produce much stress particularly if accompanied by full sun. Growing vireyas in full shade is an unsatisfactory strategy, as they do not then flower. However this problem is easily overcome with automated misting which lowers the temperature by evaporative cooling as our high temperatures occur with low relative humidity.
Of central importance to the successful cultivation of vireyas is drainage; it must be near perfect both for container culture and plants growing in the garden bed. A regular supply of water is also essential. Their water requirements are not great, compared to, say, section
Choniastrum
azaleas which have a much greater requirement, and it is preferable if between watering the plants become fairly dry at least occasionally. A well drained garden bed is easier to construct and produces better growth than container culture where good drainage requires deep pots and if these are made from black plastic, they should be kept out of direct sunlight to minimise overheating. The growing medium needs to be carefully chosen for optimum drainage, moderate water holding capacity, suitable pH, usually about 5, and an appropriate long lasting fertiliser such as Osmocote. The advantage of containers is that plants are easily moved about the garden or into heated greenhouses in regions where winters minimum air temperatures drop much below zero.
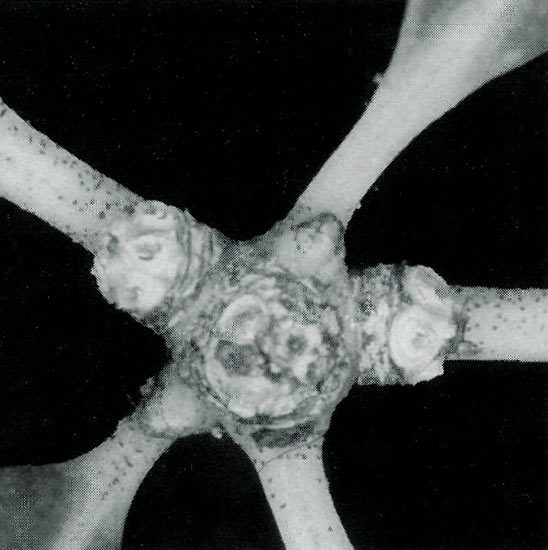
|
|
Figure 9. A stem after flowering showing three
inflorescences, one central and two lateral. |
Rhododendron loranthiflorum 'Sri Chinmoy' grows well in containers or the ground. While pots are convenient for small plants, large plants require a very large container as too small a pot inhibits growth. Over feeding should be avoided as it produces very strong growth with large internodal lengths and an absence of flowers. Usually half what is recommended on the fertiliser packet is a good starting point. This rhododendron is floriferous and under suitable conditions every terminal develops flowers. A close inspection during flowering or just after the flowers abscise shows that occasionally multiple inflorescences are produced on some terminals, e.g., a central inflorescence and one or two lateral ones (Figure 9), or lateral inflorescences with a central growth bud. Occasionally such lateral inflorescences have been seen on other vireyas, both species (e.g., R. konori , R. javanicum , R. zoelleri and R. retusum ) and hybrids (e.g., [ R. christianae x R. jasminiflorum ]) and it is likely that multiple inflorescences occur infrequently on other vireyas but go unnoticed unless the plants are carefully examined during or after flowering. Recently, Kron and Johnson (1998) reported that their evolutionary trees indicated that lateral flowers had evolved several times within Rhododendron . Our observations of occasional lateral flowers appearing in Vireya suggest that the characteristic "lateral flowers" have the potential to develop within this section.

|
| The Indian spiritual master and world peace advocate Sri Chinmoy. |
Sri Chinmoy
Sri Chinmoy is an Indian spiritual master and world peace advocate who resides in New York and has dedicated his life to showing others how to find fulfillment, happiness and peace in their lives. He believes world peace will be attained when each individual in the world experiences inner peace, which will then flow through to their outer life. He regards meditation as an important technique in achieving this inner peace by bringing to the surface the dormant peace, which lies deep within us all. For the past quarter century he has conducted twice weekly peace meditation sessions at the United Nations Headquarters in New York and he and his team of disciples have established hundreds of meditation centres around the world.
Sri Chinmoy is an author, painter, poet and musician, activities which he uses to inspire others to find their internal peace. As well, he has introduced "Sri Chinmoy Peace-Blossom Sites" in which significant places, often sites of natural beauty, are dedicated to peace and display a plaque, which encourages visitors to reflect on peace. There are many such sites round the world, and many of these are gardens since as plant lovers know, beautiful gardens induce a feeling of peace. In Australia, the National Rhododendron Garden at Olinda and some of our capital city Botanical Gardens are Peace Blossom Sites.
Today, world peace has become necessary for mankind's survival since we have more than sufficient capacity to destroy the environment of our world and all of its people, and in the event of a world war it is unlikely that this destructive technology would remain unused. It requires the efforts of many people for success in producing and maintaining world peace and it is heartening to observe that there are people who are prepared to dedicate their life to this cause. It is with much pleasure and respect that we have given the name Sri Chinmoy to this particular
Rhododendron loranthiflorum
.
Lives of great men all remind us
We can make our lives sublime,
And, departing, leave behind us
Footprints on the sands of time;
Footprints, that perhaps another,
Sailing o'er life's solemn main,
A forlorn and shipwrecked brother,
Seeing, shall take heart again.
"From "A Psalm of Life"
Henry Wadsworth Longfellow
Acknowledgments
We are greatly indebted to Sushmitam Rouse for introducing us to the guru Sri Chinmoy, acquainting us with the world wide endeavours of him and his team of disciples to establish world peace and providing us with the above information.
References
1. Bursill, L. A.; Rouse, J. L. Investigation of phyllotaxis of
Rhododendron
. In: Jean, R. V;. Barabe, D., eds. Symmetry in Plants. World Scientific Public; 1998.
2. Kron, K. A.; Johnson, S. L. Using DNA sequences to estimate evolutionary relationships among rhododendrons and azaleas. J. Amer. Rhodo. Soc. 52:70-72; 1998.
3. Rouse, J.L.; Williams, E.G. Sexual and graft compatibilities within section
Vireya
and between this section and other lepidote
Rhododendron
. Jordan, N., ed. Proc. 1994 Pacific Region International Rhododendron Conference, Burnie, Tasmania; 1997: 27-51.
4. Rouse, J.L.; Knox, R.B.; Williams, E.G. Inter- and intraspecific pollinations involving
Rhododendron
species. J. Amer. Rhodo. Soc. 47: 23-28, 40-45; 1993.
5. Sleumer, H. (1966). Ericaceae . Steenis, C.G.G.J., ed. Flora Malesiana Ser. 1 Vol. 6 (4). Noordhoff, Groningen; 469-668.
6. Stevens, P. F. The altitudinal and geo-graphical distribution of flower types in
Rhododendron
section vireya, especially in the Papuasian species, and their significance. Bot. J. Lin. Soc. 72: 1-33; 1976.
7. Stevens, P. F. Malesian vireya rhododendrons - towards an understanding of their evolution. Notes RBG Edinb. 43 (1): 63-80; 1985.
8. Williams, E.G.; Rouse, J.L. Disparate style lengths contribute to isolation of species in
Rhododendron
. Australian J. Botany 36: 183-191; 1988.
9. Williams, E.G.; Rouse, J.L. Evolutionary history and speciation in
Rhododendron
section
Vireya
. Jordon, N., ed. Proc. 1994 Pacific Region International Rhododendron Conference, Burnie, Tasmania; 1997: 52-60.
10. Wallace, A. R. The Malay Archipelago: the land of the orangutan and the bird of paradise: a narrative of travel with studies of man and nature. UK: MacMillan & Co; 1869. Reprinted 1989 by Fong & Sons Printers P/L, Singapore.