Growth Regulators in the Control of Flowering In a Vireya Rhododendron Hybrid
Richard A. Criley
Department of Horticulture
University of Hawaii
Honolulu, HI
Abstract
Native to the mountain forests of southeast Asia and the Indo-Pacific islands, vireya rhododendrons are evergreen and have a wide array of flower shapes and colors, with several species having fragrance. Their growth habit tends to be lanky, and flowering times are uncertain. Under a grant from the American Rhododendron Society, experiments were conducted on a unnamed hybrid of
R. aurigeranum
x
R. herzogii
to find ways to produce compact potted plants and to control flowering. Synthetic cytokinins* (200 ppm) applied as foliar sprays to the foliage of branches pruned back to a whorl of leaves stimulated 94 to 96% bud break, versus 88% for pruned control plants. As with temperate rhododendrons, most of the growth retardants** were effective in controlling elongation growth as drenches, while sprays were less effective. The growth retardant paclobutrazol (PBZ) is recommended as a drench of 1 to 2 mg per 15 cm pot to control growth. Anthesis (flower opening) was reached 21 to 26 weeks after treatment with PBZ.
Introduction
Rhododendrons of the section
Vireya
of the genus have been cultured for more than 150 years in botanic gardens, commercial nurseries, and collections of hobbyists (Cavender, 1994; Kenyon and Walker, 1997; Leach, 1987). Plant breeders have developed hybrids that are often more vigorous and floriferous than the original species. This vigor, however, lends itself better to their use as landscape plants than potted plants, as the stem length between whorls can be 10to 20 cm long. While some species have well-defined flowering periods, many hybrids have long flowering seasons. In Hawaii, some vireya rhododendrons have been found in bloom nine months out of the year (Bob Hirano, Lyon Arboretum, personal communication). Flowering times are uncertain and this feature was one of the focuses of this research, together with the control of their somewhat lanky growth for potted plant culture.
Vegetative growth of azaleas and rhododendrons may be controlled using growth retardants, and flower bud initiation is often a result of applications of these compounds (Cathey, 1965; Criley, 1985; Gent, 1995; Larson, 1995; Ranney, etal., 1994; Stuart, 1961; Wilkinson and Richards, 1991). Application of these retardants to the vireya rhododendrons appeared likely to have similar results.
Synthetic cytokinins stimulate bud break in ornamentals and fruits (Parups, 1971; Carpenter, etal., 1971; Greene and Autio, 1990; Henny and Fooshee, 1990; Keever and Foster, 1990; Quinlan and Tobutt, 1990; Williams and Stahly, 1968). Such reports suggested the use of synthetic cytokinins to improve the branching of the vireya rhododendrons.
Under a grant from the American Rhododendron Society, the objectives of this work were to induce better branching, to increase plant compactness, and to define a protocol that would permit controlled flowering in potted vireya rhododendrons. Success in this research could lead to increased production of these attractive plants for the florist and garden center trade.

|
Materials and Methods
The plant materials used in these experiments were an unnamed
R. aurigeranum
X
R. herzogii
hybrid and its reciprocal (
R. herzogii
X
R. aurigeranum
) (R. Hirano, Lyon Arboretum). The plants were grown from rooted cuttings in 15 cm pots in a medium of equal parts coarse bark, coarse sphagnum peat moss, redwood compost, and volcanic cinders amended with controlled release fertilizer (Osmocote 14N-6P-11.6K), dolomite, and superphosphate at the rates of 1, 1.5, and 1.5 kg/cu. m respectively. The plants were treated after three to five flushes of growth had developed. The environmental conditions were 30% or 50% shade under woven polypropylene, ambient temperatures (range of 17 to 27°C, depending upon season), and daily irrigation by spray stakes.
In the first experiment, to induce more lateral breaks, synthetic cytokinins were applied to pinched and un-pinched plants by foliar spray to both upper and lower leaf surfaces to thorough wetness. The cytokinins (N-6-BA, PBA, TDZ) were prepared by diluting acetone stock solutions with water or by dissolving crystals in dimethylsulfoxide (DMSO) and diluting to 2.5% DMSO to achieve active ingredient concentrations of 200 mg/L. A wetting agent (Tween 20) was added to achieve a concentration of 0.5%.

|
|
Treatment with 200 ppm of the cytokinin PB
increases lateral bud break on pruned shoots. |
In the second experiment, growth retardants (paclobutrazol, uniconazole, chlormequat, flurprimidol, and daminozide) were prepared by diluting commercial stock solutions to the desired active ingredient concentrations. These concentrations were based upon successful responses of temperate rhododendron species as reported by Larson (1995), Ranney etal. (1994), and Wilkinson and Richards (1991). Drenches of paclobutrazol, uniconazole, chlormequat and flurprimidol were applied in volumes of 100 ml/15 cm pot. Daminozide was applied as a foliar spray to run-off to both upper and lower foliage surfaces.
The plant material used in a third experiment was the reciprocal cross, R. herzogii X R. aurigeranum (R. Hirano, Lyon Arboretum). This experiment combined the use of 200 mg/L PBA applied to all plants with PBZ drenches of 0, 1, 2 or 4 mg/15 cm pot 5 weeks later when new shoots were 2 to 3cm long. After two months growth under 30% saran shade, the plants were placed in controlled temperature growth chambers (22/17, day/night; irradiance of 360 watts per sq. meter from cool white fluorescent lamps) with day lengths of either 11 or 16 hours.

|
|
Unnamed
R. aurigeranum
x
R. herzogii
(R. Hirano, Lyon Arboretum)
remains compact with high bud count following treatment with the growth retardant paclobutrazol. |
In the fourth experiment, cuttings were potted three per pot and the tip of new growth was pinched after the first flush from the cutting had completed elongation. At the time of retardant treatment there was a long flush of one to several branches. These were not pinched again. Because a previous experiment had suggested that additional phosphorus would improve flower bud set, some plants received three drenches of a liquid formulation of a high phosphorus fertilizer (4-30-8, Nutri-Grow, San Diego, CA) at the rate of 1 ml/200 ml water. Phosphorus in this formulation is in the phosphite form. The treatments were (1) control, (2) three fertilizer drenches 10 days apart, (3) paclobutrazol (PBZ) at 2 mg active ingredient per 15 cm pot, (4) a combination of treatments 2 and 3, and (5) a repeat of treatment 4 initiated four weeks after treatment 4.
In each experiment, treatment units consisted of five to eight plants. Each plant was treated as a single replicate, and plants were randomized on the bench. In the bud break experiment, shoots were cut back to a strong whorl of leaves with no lateral shoot development, and the number of potential bud break sites was determined by counting leaves. In growth retardant experiments, three to five terminals were marked with paper tags so that elongation growth from the last whorl present at time of treatment could be measured. Flower bud counts and days to flower were also recorded. Plant heights were measured from the pot rim to the plane of the topmost whorl of leaves.
Results
Lateral bud break on cytokinin-treated plants began about three weeks after pruning and cytokinin treatment. Shoot counts taken eight weeks after treatment showed that 200 mg/L of PBA or TDZ stimulated 96.4% and 94.5% (respectively) of the axillary buds to develop as shoots, while on the 200 mg/L N-6-BA and the pruned-but-untreated control plants 88.1% bud break occurred. Un-pruned controls had 63% bud break at whorls located at the same level as pruned plants. Elongating shoots from TDZ treated whorls were shorter than shoots from other treatments. The next flush of shoots from those arising at cytokinin-treated whorls did not show an increased tendency to branch.
| TABLE 1. Response of the hybrid R. aurigeranum X R. herzogii to application of growth retarding compounds. | ||||||
| Retardant |
Rate
S=mg/L D=mg/pot |
Appln
Method |
Elongation (cm) of youngest flush |
No.
Budded plants |
Days from
PGR Trt to flower |
|
| 8 wk | 15 wk | |||||
| Control | 8.8 | 2.3 | 2 | 140 | ||
| Chlormequat | 1000 | Drench | 4.2 | 3.2 | 8 | 143.1 |
| Daminozide | 5000 | Spray | 6.5 | 3.6 | 5 | 146.4 |
| Flurprimidol | 2 | Drench | 5.0 | 3.7 | 8 | 158.7 |
| Paclobutrazol | 4 | Drench | 3.3 | 0 | 7 | 122.3 |
| Uniconazole | 4 | Drench | 1.4 | 0 | 8 | 144.2 |
| Treatment effects | 0.05 | 0.05 | NS | |||
| BLSD 0.05 | 2.73 | 1.70 | -- | |||
| Bayesian Least Significant Difference: Differences between any 2 means in a column that exceed the BLSD value are significant at the 0.05 level. | ||||||
| TABLE 2. Response of the hybrid R. aurigeranum X R. herzogii to drench applications of paclobutrazol (2 mg/plant) and/or phosphorus fertilization (3 applications 10 days apart of 4-30-8 soluble fertilizer diluted at 1:200 ml). Treatments initiated on May 3 except last paclobutrazol + phosphorus treatment on May 30. | ||||||
| Treatment | Percentage of flowering plants | Mean No. flower buds per plant | Average date of first flowering | Days from treatment to first flower | Average date of peak bloom |
Mean plant height
(cm) |
| Control | 0 | -- | -- | -- | -- | 47.7 |
| Phosphorus | 14 | 2 | Oct. 3 | 153 | Oct. 5 | 34.9 |
| Paclobutrazol | 100 | 6.8 | Sep.29 | 149.4 | Oct. 8 | 13.4 |
| Paclobutrazol + phosphorus | 100 | 8.7 | Oct. 2 | 152.1 | Oct. 9 | 12.4 |
| Paclobutrazol + phosphorus (4 week delay) | 43 | 8.3 | Nov. 4 | 157 | Nov. 7 | 18.4 |
| LSD (0.05) | 1.28 | 2.62 | ||||
All retardant-treated plants had shorter growth flushes than did the control (Table 1) and some produced fewer flushes of growth before initiating a flower bud. High percentages of budded plants were produced by drench treatments of flurprimidol, chlormequat, uniconazole, and paclobutrazol, while only 25% of control plants set bud and flowered. Chlormequat-treated plants showed the typical chlorosis following treatment. The earliest evidence of flower bud formation was at eight weeks after treatment with the PBZ drench, and all retardant treatments had some plants with buds at fifteen weeks. The PBZ-treated plants flowered earliest, averaging 122 days after treatment, while flurprimidol-treated plants averaged 158.7 days to anthesis following treatment.
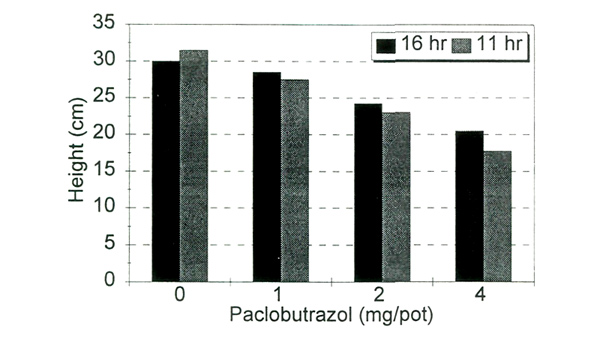
|
| Fig. 1. Plant height as a response to paclobutrazol treatment and day length. |
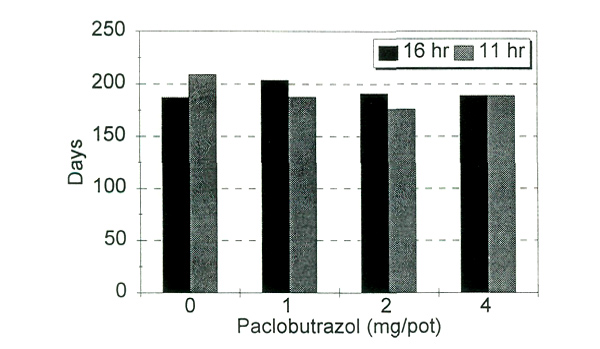
In the third experiment, optimum rates of PBZ were 1 and 2 mg/15 cm pot as the higher rate permitted almost no shoot elongation. Inflorescence initiation began about eight weeks after the PBZ drench, and anthesis ranged from 176 to 209 days. As retardant concentration increased, the overall plant height was shorter (Fig. 1), the percentage of terminals setting flower buds increased (Fig. 2), but days to flower did not differ significantly (Fig. 3). The day lengths to which plants were exposed during initiation and development did not significantly affect time to bloom (Fig. 3), but there was a slight increase in bud count under 16-hour days over all retardant treatments.
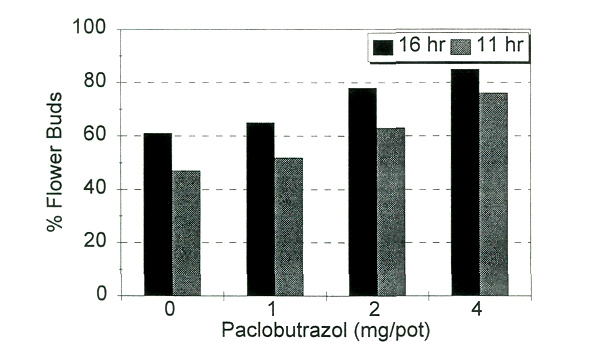
|
|
|
|
Fig. 2. Flower buds as a percentage of total shoot number as affected
by paclobutrazol treatment and day length. |
Fig. 3. Days to flower from paclobutrazol treatment as influenced
by paclobutrazol treatment and day length. |
In the fourth experiment, plants receiving no retardants (control) did not set flower buds by the time that PBZ-treated plants had budded and flowered (Table 2). Only one plant of the high phosphorus-only plants had set a flower bud, and this plant flowered at about the same time as the PBZ and PBZ + phosphorus plants flowered. Anthesis was reached about five months after paclobutrazol treatment for PBZ alone and PBZ + high phosphorus fertilizer. Flowering of plants in the 4-week delay PBZ + high phosphorus treatment had basically the same five month response period as the plants that were treated four weeks earlier. Three flushes of growth developed on control and phosphorus-only plants while only one very short flush developed following paclobutrazol treatment. Instead of another vegetative flush, an inflorescence bud was initiated.

|
|
An untreated plant (left) was taller and set fewer
flower buds than a paclobutrazol-treated (4 mg/pot applied as a drench) plant. |
Discussion
Increased lateral branching from pinched or pruned stems results from the application of cytokinin in comparison to plants that have not been pinched. This imparts a more compact appearance to the potted plant. While PBA was superior in this experiment, N-6-BA may be as effective if a higher concentration is applied. TDZ is not yet available in formulations suitable for use as a branching inducer. Subsequent growth flushes did not show increased lateral branching due to the application of the synthetic cytokinins.
The vireya hybrid used in experiment three did not show a marked response to 11 - and 16-hour photoperiods during flower initiation and development. While azaleas seem to initiate flowers sooner on short photoperiods (Criley, 1969a) and some of the temperate rhododendrons initiate flowers on long days (Cathey, 1965; Criley, 1969b), this representative of the genus appears to be less responsive.
As reported for other rhododendrons (Larson, 1995; Wilkinson and Richards, 1991) and mountain laurel (Banko and Bir, 1999), shorter stems and improved flower bud set in these two vireya hybrids resulted from the application of growth retardants. The broader labelling of PBZ, in comparison to other retardants, makes it the retardant of choice to achieve compact potted rhododendrons. While PBZ at 2 mg active ingredient per 15 cm pot was an effective retardant, it was excessive on some of the other species tested (data not reported). Over-treatment is possible when it is used as a drench; thus experimentation will be necessary to determine optimum rates to use in different media and with different species. At the 1 to 2mg application rate, PBZ shortened growth of the next one or two growth flushes on potted plants following flowering before normal elongation resumed. Plants at the 4 mg rate had not begun to grow normally a year after treatment.
Reports of enhanced flower bud formation in field-grown temperate rhododendrons with phosphorus fertilization (Myhre and Moretensen, 1964) suggested the application of phosphorus to the vireyas. Addition of phosphorus to the potting medium via liquid feed did not make a significant difference in the number of flower buds set in this experiment although the trend of 1½ to 2 buds additional over the PBZ-only treatment was interesting.

|
|
The growth retardant paclobutrazol as a 2 mg/pot drench increases
flower bud set on the hybrid R. aurigeranum x R. herzogii . |
The flower bud-inducing properties of PBZ and other retardants enhance the floral display properties of the final product. In these experiments with the limited range of the two hybrids, flowering was achieved at five to six months after drench treatment with PBZ. The timing and scheduling of retardant applications remain to be worked out for different species and cultivars, as demonstrated by the difference in response and development time for the two hybrids used in these experiments. This report may stimulate trials on the part of commercial potted plant producers and curious vireya growers.
* Synthetic cytokinins are chemicals similar to naturally-occurring plant hormones that stimulate bud-break and branching. The cytokinins used in this experiment were N-6-benzylaminopurine (N-6-BA) from Abbott Labs; N-(phenylmethyl)-9-(tetra-hydro-2H-pyran-2yl)-9h-purin-6-amine (PBA) from Abbott Labs; and N-phenyl-N'-1,2,3-thiadiazol-5-ylurea (thidiazuron, TDZ), from AgroEvo.
** Growth retardants are chemicals that reduce the production of the natural stem-elongating hormones of plants and cause shorter growth flushes. In these experiments, the growth retardants included Cycocel (chlormequat chloride), a product of American Cyanamid, now marketed by Olympic Chemical; B-nine SP (daminozide), a product of Uniroyal Chemical; Bonzi (paclobutrazol), a product marketed by Uniroyal Chemical; A-rest (ancymidol), a product of Dow Elanco now marketed by Sepro Corp.; Cutless (flurprimidol) marketed by Dow AgroSciences; and Sumagic (uniconazole), marketed in the U.S. by Valent Chemical Co.
Acknowledgment
This research was supported, in part, by a grant from the American Rhododendron Society. The author is Professor of Horticulture at the University of Hawaii, Honolulu, HI. Published as Journal Series 4453 of the College of Tropical Agriculture and Human Resources, University of Hawaii.
Literature Cited
1. Banko, T. J. and R. E. Bir. Use of growth retardants to promote flowering of Mountain Laurel,
Kalmia latifolia
L. J. Environ. Hort. 17:11-17; 1999.
2. Carpenter, W. J., R C. Rodriguez, and W. H. Carlson. Growth regulator induced branching of non-pinched poinsettias. HortScience 6:457-458; 1971.
3. Cathy, H.M. Initiation and flowering of
Rhododendron
following regulation by light and growth retardants. Proc. Amer. Soc. Hort. Sci. 86:753-760; 1965.
4. Cavender, R. Rare beauties. Interior Landsc. 11 (2):10-1 7; 1994.
5. Criley, R. A. Controlling rhododendron flower bud initiation. Flor. Rev. 143(3715):36-37, 93-95; 1969a.
6. Criley, R. A. Effect of short photoperiods, Cycocel, and gibberellic acid upon flower bud initiation and development in Azalea 'Hexe'. J. Amer. Soc. Hort.Sci. 94:392-396; 1969b.
7. Criley, R. A. Rhododendrons and azaleas. Handbook of Flowering 4:180-197. CRC Press, Boca Raton, FL; 1985.
8. Gent, M. P. N. Flowering of
Rhododendron
and
Kalmia
in response to application date of Bonzi or Sumagic. Proc. Int. Plant Prop. Soc. 45:543-545; 1995.
9. Greene, D. W. and W. R. Autio. Vegetative responses of apple trees following benzyladenine and growth regulator sprays. J. Amer. Hort. Sci. 115:400-404; 1990.
10. Henny, R. J. and W. C. Fooshee. Thidiazuron stimulates basal bud and shoot formation in
Alocasia
X
Chantrieri
Andre. HortScience 25:124; 1990.
11. Keever, G. J. and W. J. Foster. Chemically induced branching of woody landscape plants. J. Environ. Hort. 8:78-82; 1990.
12. Kenyon, J. and J. Walker. Vireyas - A practical gardening guide. Timber Press, Portland, OR; 1997.
13. Larson, P. C. Use of paclobutrazol to regulate flower bud initiation and stem elongation in
Rhododendron catawbiense
'Boursault' Proc. Int. Plant Prop. Soc. 45:459-463; 1995.
14. Leach, D. Vireya rhododendrons. Horticulture 65:30-35; 1987.
15. Myhre, A. S. and W. P. Mortensen. The effect of phosphorus on rhododendron flower bud formation. Amer. Rhodo. Soc. Quart. Bull. 18(2):66-71; 1964.
16. Quinlan, J. D. and K. R. Tobutt. Manipulating fruit tree structure chemically and genetically for improved performance. HortScience 25:60-64; 1990.
17. Parups, E. V. Use of 6-benzylamino purine and adenine to induce bottom breaks in greenhouse roses. HortScience 6:456-457; 1971.
18. Ranney, T. G., R. E. Bir, J. L. Conner, and E. P. Whitman. Use of paclobutrazol to regulate shoot growth and flower development of 'Roseum Elegans' rhododendron. J. Environ. Hort. 12:174-178; 1994.
19. Stuart, N. Initiation of flower buds in Rhododendron after application of growth retardants. Science 134:50-52; 1961.
20. Wilkinson, R.I. and D. Richards. Influence of paclobutrazol on growth and flowering of
Rhododendron
'Sir Robert Peel'. HortScience 26:282-284; 1991.
21. Williams, M. W. and E. A. Stahly. Effect of cytokinins on apple shoot development from axillary buds. HortScience 3:68-69; 1968.
Synopsis
The open and somewhat lanky growth habits of vireya rhododendrons have limited their usage as potted plants among the garden center and florist trades. A grant from the American Rhododendron Society supported research aimed at developing more compact plants with good floral displays for holidays, shows, and regularized marketing.
To this end, chemicals, known as growth retardants, which shorten the elongation of growth flushes, and synthetic compounds (synthetic cytokinins) that mimic the natural branching stimulators of plants were applied to an unnamed hybrid of Rhododendron aurigeranum X R. herzogii and its reciprocal cross. The research was conducted on potted plants at the University of Hawaii in Honolulu, Hawaii. The results showed that the foliar application of synthetic cytokinins could increase the number of lateral branches that result following a pinch. Several growth retardants could increase compactness by shortening growth flushes, as well as increase flower bud production. The retardant known by its trade name of Bonzi (paclobutrazol is the active ingredient) is recommended for use as a drench to the medium at the rate of 1 to 2 mg active ingredient per 15 cm (6-inch) pot.
[The commercial formulations of synthetic cytokinins are either not available in the U.S. or are not registered for use as branching stimulators on rhododendrons. The synthetic cytokinin N-6-BA can be purchased from various chemical supply companies (for example, Sigma-Aldrich, www.sigma-aldrich.com). The growth retardants are normally sold by horticultural supply companies and can sometimes be found at the larger garden centers. For Bonzi (paclobutrazol), check: www.dowagro.com or www.uniroyalchemical.com for suppliers or purchasing information.]