Taxonomy of the Flowering Forms of
Rhododendron calendulaceum (Michx.) Torrey
Frank F. Willingham, Jr., Ph.D., Pine Mountain, Georgia
In 1973, I reported on the initiation and current status of my taxonomic study being conducted throughout the Nantahala Mountain region of Southwestern North Carolina and northern Georgia (Willingham, 1973). For some time, this area has been noted for its unusual and extremely variable native azaleas( especially
R. calendulaceum
(Braun, 1941). It was after collecting in this region that Dr. Henry Skinner (1955) suggested the existence of a distinct third phase or bloom form of
R. calendulaceum
, possibly composed of
R. calendulaceum
x
bakeri
hybrids and backcrosses. The existence of the other two phases or forms of the species was evidently well established and they have been described on several occasions in the following way (Galle, 1968; Skinner, 1961)
Early-Flowering
R. calendulaceum
- the "typical" species, bearing large, yellow to orange flowers (averaging 48 mm. across wing petals), occurring at low altitudes (below 925 m.), and flowering before the leaves are mature and expanded (usually late April to early June).
Late-Flowering
R. calendulaceum
- usually a smaller, more stoloniferous plant, bearing smaller red or reddish-orange flowers (averaging 42 mm. across wing petals), occurring at higher altitudes (above 925 in., and flowering after the leaves are fully mature (usually June to mid-July).
To the taxonomist, a question arises as to what is meant by the terms "phase" and "form". In the species concept of
R. calendulaceum
as it seems to be understood by most horticulturists and azalea enthusiasts I have known, phase and form are like subcategories of a species. In other wards, they carry a definite taxonomic connotation; they are easily recognizable categories, that have a status just below the rank of species. To my knowledge, however, no one has conducted a detailed study of just the single species
R. calendulaceum
over its entire altitude range and flowering season to determine whether our notions about the flowering forms are correct. This is what I did. In addition, I was interested in the role
R. bakeri
has played in contributing to the apparent uniqueness of the Nantahala azaleas, and therefore sought to determine its relative abundance and possible involvement as a hybrid parent in that area.
Methods
In order to compare the flowering forms of
R. calendulaceum
, three types of data were collected: (1) morphological, (2) chemical, and (3) cytological. Morphological data were obtained by screening over 500 individual plants for ten characters. Each character was quantified and the numerical values obtained from the collections were analyzed statistically by computer. The characters used included (1) leaf maturity at time of flowering, (2) predominant flower color, (3) corolla width, (4) pollen size, (5) percent pollen stain-ability, (6) degree of lower cuticular ridging, (7) stomate size, (8) stomate density, (9) presence of a color blotch on the upper petal, and (10) growth habit.
Cytological data included only chromosome numbers obtained from cuttings rooted under mist. No meiotic figures were observed.
Chemical data consisted of thin-layer chromatographic patterns of leaf flavonoids. The extraction and hydrolysis of leaf flavonoids to give the aglyconemoity followed the pattern of Bate-Smith (1956, 1958). Thin-layer plates were coated with microcrystalline cellulose and run one direction in Forestal Solvent after the method of Harbone and Williams (1971). The dried plates were then viewed in the presence of ammonia vapor under long-wave ultraviolet light.
|
|
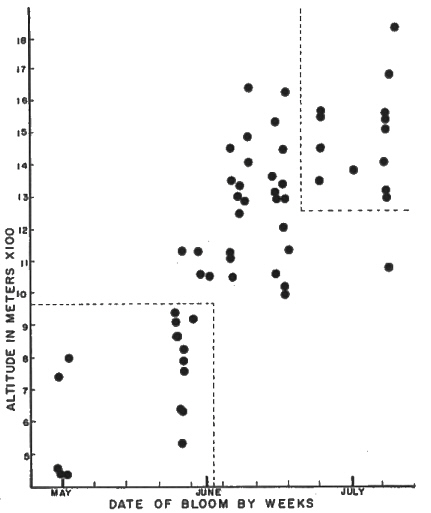
FIG. 37. Date vs. Altitude for all azalea collections. The dotted
lines indicate the collections which were arbitrarily assigned as early-flowering form or late-flowering form. Note that there is no overlap between the groups. |
Results and Conclusions
Reference to Fig. 37 will show that the flowering pattern in
R. calendulaceum
, when followed continuously through the spring and summer, is different from that described by Dr. Skinner (1955). No distinct phases were obvious and the general impression was simply that as the summer progressed, plants came into flower higher on the mountains. This is typical of many Southern Appalachian species with ranges extending over large altitude gradients. In order to facilitate comparisons between the now questionable flowering forms or phases, several populations were arbitrarily assigned early-flowering and late flowering status as indicated by the dotted lines on Fig. 37.
Analysis of the morphological characters resulted in a wealth of numbers which I have not reproduced here, but which are available if requested. Summarizing these data, it was found that if only the means are considered, there is a statistically significant difference between the arbitrarily assigned flowering forms in terms of leaf development at flowering time, corolla width, presence of a blotch on the upper petal, cuticular pattern, stomate size, and percent pollen stainability. If, however, the range and degree of character variation within each group are considered, there is so much overlap that it becomes impossible to distinguish between the flowering forms on the basis of characters used. Put in practical terms, if I gave you a single plant, I believe it would be impossible for you to tell me with certainty whether it was an early-flowering form or a late-flowering form. Such distinctions must be clear if the forms or phases are to have taxonomic meaning.
Neither the chromatographic nor cytological data provided any convincing evidence for differences in the so-called flowering forms. Thin-layer chromatographic patterns were virtually identical for plants tested from both arbitrarily selected groups and only normal tetraploid (2n=52) chromosome numbers were encountered in any of these plants. The conclusion, then, is that there really are not any recognizable, distinct phases or flowering forms within the species
R. calendulaceum
, at least not taxonomically significant ones.
R. calendulaceum
, like other of the native species, is highly variable.
It is not unusual to encounter in the field a population of what appear to be significantly distinct individuals. I believe that there is considerable danger in rushing to name new forms, new varieties, or any new subspecific taxa without a thorough investigation of their biological reality. To me, the better choice is to accept the species concept as a broad one which may include many unusual and variable individuals. After all, that's what we do with people!
|
|
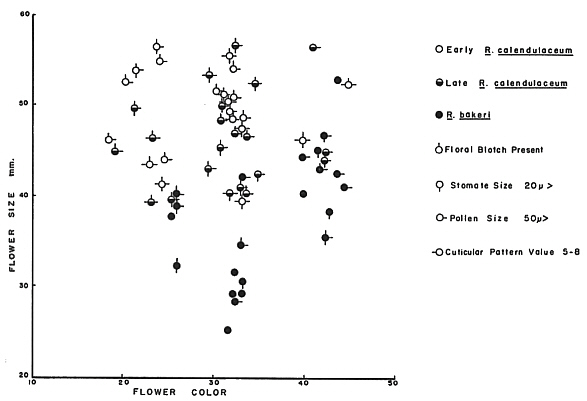
FIG. 38. Pictorialized scatter diagram illustrating the combined variation in nine
characters for native azalea collections. |
What about
R. bakeri
? The first problem I encountered with this species was extreme difficulty in locating and positively identifying it. Apparently it is much more distinct in its Virginia and Kentucky habitats than in North Carolina and northern Georgia. In the latter two it is easily confused with
R. calendulaceum
by virtue of great overlap in practically every morphological character. The only way I was eventually able to positively identify the species was by chromosome count (2n=26) or chromatographic pattern. Both techniques are unsuitable for field identifications, and therefore since good "clean" diagnostic characters seem to be lacking, a review of the taxonomic rank of this plant would seem in order. No triploid chromosome numbers, suggestive of
R. calendulaceum
x
bakeri
hybrids, were ever encountered nor did any of the chromatographic data indicate the presence of such hybrids among my collections. Figure 38, a pictorialized scatter diagram which simultaneously plots the relationships of nine separate characters, shows clearly the lack of distinctiveness among the three entities under consideration. Trends may be seen, but no clean breaks, which are needed to provide the discontinuities necessary for recognition and taxonomic separation.
I have concluded that
R. bakeri
is also a highly variable species, not well separated from
R. calendulaceum
in the southern parts of its range. It needs to be studied closely, perhaps with the idea that it is a cytological race of
R. calendulaceum
. I have found no conclusive evidence for its having hybridized extensively with
R. calendulaceum
and I would urge caution in reaching such conclusions solely on the basis of morphological characters. The Gregory Bald azaleas have shown that it is not always possible to tell the parents just by looking at the offspring (Li, 1957).
One interesting surprise which has resulted from this study is the finding that pollen fertility is much lower in
R. bakeri
than in
R. calendulaceum
. I have also noted that our plants of
R. bakeri
at Callaway Gardens do not set seed well, and one wonders at the apparent success of this species (?) under such a competitive disadvantage. This knowledge has prompted a new and detailed investigation of pollen in the native azaleas, about which I hope to report in the near future.
References
- Bate-Smith, E. C., 1956, The commoner phenolic constituents of plants and their systematic distribution, Sci. Proc. Royal Dublin Soc., 27: 165-176
- ____ 1958, The phenolic constituents of plants and their taxonomic significance, J. Linn. Soc. (Bot.), 58: 371-395.
- Galle, Fred C., 1968, Native and Some Introduced Azaleas for Southern Gardens, Booklet No. 2, Callaway Gardens, Pine Mountain, Ga. 28 ppg.
- Harborne, J. B. and Christine A. Williams, 1971, Leaf survey of flavonoids and simple phenols in the genus Rhododendron, Phytochem, 10: 2727-2744
- Li, Hui-Lin, 1957, Chromosome studies in the azaleas of eastern North America, Am, J. Bot., 44:8-14.
- Skinner, Henry T., 1955, In search of native azaleas, Morris Arboretum Bulletin, 6:3-10, 15-22.
- ____ 1961, Classification of the native American azaleas, Proc. Int. Rhododendron Conference, Portland, Oregon, The Amer. Rhod. Soc., pgs. 81-86.
- Williamgham, Frank F., Jr., 1973, Variation in Rhododendron calendulaceum, a return to the Nantahala, Bull. Amer. Rhododendron Soc., 27(2): 117-118.