Breeding For The Heat Resistant Rhododendrons
Part II: Differences Among Species And Hybrids
Ken-ichi Arisumi, Eisuke Matsuo, Yusuke Sakata and Tsutomu Tottoribe
Laboratory of Ornamental Horticulture and Floriculture
Kagoshima University, Kagoshima, Japan
Reprinted from The Memoirs of The Faculty of Agriculture, Kagoshima University
Vol. XIX (Whole Number 28), March 1983, Kagoshima, Japan.
Editor's note: Part I appeared in the ARS Journal, Vol. 39:4, Fall 1985.
Introduction
The distribution of elepidote rhododendrons is almost entirely confined to the wetter, high mountain regions. The centre of origin is said to be in the area where Tibet, Burma and Yunnan meet. The areas ranging from the Himalayas through eastern Tibet, upper Burma to western and central China have been inhabited by a great number of species.
From these areas they have extended their distribution, eastwards to Taiwan, north-eastwards through Japan and Siberia to the north-western and north-eastern United States, westwards to the Caucasus and the Pyrenees, or southwards to Ceylon or Malaya and Sumatra. In the mountain regions of these areas blest with cool summers, sporadic species are found.
Moreover, the hybridizations to create the garden forms have been mainly conducted in the areas with cool summers, i.e., in north-western Europe and the north-eastern or north-western United States.
Therefore, it has been very difficult to grow rhododendrons successfully in the lowlands of the southwestern part of Japan; almost all of the natural species and their garden hybrids of foreign origin have never given good results in this region where the summer is exceedingly hot and radical changes between rainy and sun scorching weather prevail.
In the previous paper, (1) the hybrid seedlings derived from the cross between 'Ruby Hart' and
R. metternichii
were revealed to grow more vigorously, tolerating more heat than either parent and a major cause was attributed to heterosis.
The present investigation was conducted to get some clues to breeding for heat resistant rhododendrons from the comparisons of the growth-differences in the seedlings of various species and hybrids.
Materials and Methods
Seeds used were chiefly obtained from the Seed Exchange System of the Japanese Rhododendron Society and H.L. Larson Rhododendrons of the U.S.A. Some seeds were also obtained from Yoshioka Nursery and the other seeds such as;
R. yakushimanum
x
R. simiarum
and
R. yakushimanum
x (
R. yakushimanum
x
R. wrayi
) were due to the courtesy of Dr. P.G. Valder of Sydney University;
R. metternichii
var.
hondoense
f.
brevifolium
and its hybrid with 'Gil's Crimson' were from Mr. T. Nagami;
R. wrayi
was from our collection at Malaya.
Seeds were sown from late January to early February in 1980 and the seedlings obtained were transplanted into 36 x 45 x 10 cm plastic boxes from April 20 to May 3, 1980. The media used was a mixture of fine pumice, peat and shredded sphagnum moss, 6:3:1 by volume. They were grown in a vinyl-house covered with black lawn, two layers in 1980 and one layer in 1981. Liquid fertilizer containing N, P and K, 50, 25 and 40 ppm, was given every 3-4 days in the growing season of 1980 and that containing 100, 50 and 80 ppm was given every 7 days in 1981. In the majority of cases the number of individuals in each seedling group at the start of the experiment was 150 plantlets, but ranged from some hundreds in the promising species or hybrids to some tens for those which we could not obtain sufficient seeds.
The mean maximum and minimum temperatures in the vinyl-house in the summer half of 1981 were as follows; May, 34.4 and 13.4°C; June, 36.9 and 18.9°C; July, 40.1 and 24.8°C; August, 38.7 and 23.6°C; September, 34.0 and 20.6°C and October, 31.0 and 14.7°C, respectively.
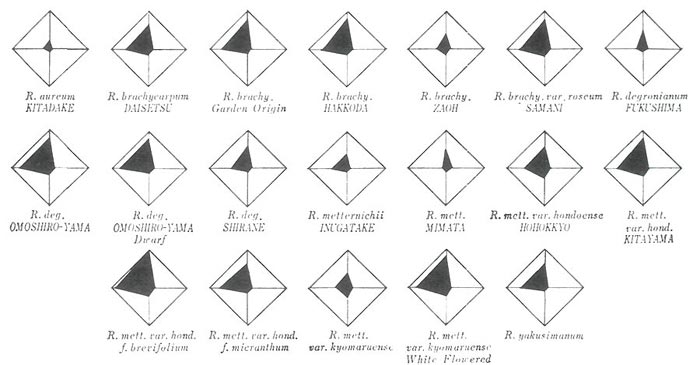
The measurements of seedling growth were done every 45 days in both the growing seasons, but for plant survival only the final data of Nov. 20, 1981, and for vigour, plant height and leaf size those of Jan. 4-7, 1982, were summarized and presented in Figs. 1-3. In these figures each corner of square was survival ratio (left), vigour (top), plant height (right) and leaf size (bottom), respectively. The survival ratio was the percentage of individuals that finally survived. The degree of vigour was based on a five grade estimation from ±, through +, ++, +++ to ++++. The plant height was measured of the largest 30 individuals selected within each seedling group and the leaf size was based on the measurement of length and width of the largest leaf in each individual of these 30 plantlets selected. In Figs. 1-3, both measurements were presented on the percentage basis to the largest seedling group, i.e., in the plant height the percentage to the value of 'Purple Splendour' x 'Scandinavia', and in the leaf size to the value of 'Blue Peter' x 'Scandinavia'.

|
|
Fig. 1. The survival ratio, vigour and growth of the seedlings in the species of foreign origin. Each
corner of square was survival ratio (left), vigour (top), plant height (right) and leaf size (bottom), respectively. |
Results and Discussion
1. Species of foreign origin
As seen in Fig. 1, there occurred a considerable difference in the heat resistance between series, sub-series and species. A large number of species displayed poor behaviour for use as the cross parent for future breeding. Especially in
R. auritum
of Boothii series,
R. haemaleum
of Neriiflorum series and
R. wardii
of Thomsonii series, all plantlets died. However, the species belonging to Fortunei series, subseries Fortunei gave fairly consistently good results as seen in
R. chlorops,
R. decorum
,
R. diaprepes
,
R. discolor
and
R. fortunei
, although no good results were obtained in
R. houlstonii
belonging to the same subseries and in the following species belonging to the other subseries of the same series, i.e., in
R. sutchuenense
of subseries Davidii, in
R. cardiobasis
and
R. orbiculare
of subseries Orbiculare and in
R. oreodoxa
of subseries Oreodoxa. The results of
R. praevernum
of subseries Davidii and
R. griffithianum
of subseries Griffithianum were intermediate.
In others,
R. elliottii
of Irroratum series and
R. ponticum
of Ponticum series gave satisfactory results. However,
R. wrayi
of Irroratum series whose habitat is in the mountain regions of Malaya and, thus, being one of the most southern species among the elepidotes, displayed unexpectedly poor behaviour, probably because it could not readily prevail over the water-logged state of the growing medium.
Of particular interest was
R. formosanum
, which is distributed in the mountain regions of middle altitudes throughout Taiwan (5). This species belongs to the Arboreum series, subseries Argyrophyllum. From the distribution pattern of various species within this subseries, the centre of origin of Argyrophyllum is assumed to be in west Szechuan. Therefore,
R. formosanum
is one of the most remote, marginal species.
The struggle in the evolutionary processes during species formation, in other words, the emulation in the dispersal processes from the high mountains of Szechuan towards the east to Taiwan, might afford this species a sort of capacity to tolerate unfavourable circumstances. This would also be the reason why the distribution of this rhododendron could remain in broad-leaved forests at the middle altitudes throughout the island and pass off without climbing up to the tops of the mountains.
Similar but somewhat inferior results were obtained with
R. morii
and
R. pseudochrysanthum
of the Barbatum series of Taiwan origin. When compared with
R. formosanum
, it would be of interest that the species from higher altitudes gave consistently more inferior results. However, the results of these two species were superior to those of
R. barbatum
and
R. strigillosum
. As in the case of
R. formosanum
, this would indicate the similar acquisition of tolerance by these two species during their geographical dispersal, although this phenomenon is not necessarily a general rule as seen in
R. wrayi
,
R. aureum
,
R. catawbiense
and
R. maximum
.
As compared with the results of its hybrids, mentioned later,
R. hyperythrum
didn't give as good a result as initially expected. The reason is not clear. The seed source of this species was from the United States, accordingly further investigations with other seed sources will be needed.
Hoitink etal. (3, 4) studied the resistance of various rhododendrons to root rot caused by
Phytophthora cinnamomi
, the most serious disease of rhododendrons, and revealed that the majority of species and cultivars are highly susceptible and can succumb to this disease. The fungus is furious in soils of high temperature with high water content. Thus, it will be practically impossible to exclude the resistance to this fungus from the breeding for heat resistant rhododendrons in the south-western part of Japan.
Although Hoitink etal. (3, 4) did not study the resistance of
R. morii
and
R. formosanum
to this fungus, they evaluated
R. pseudochrysanthum
and
R. hyperythrum
to be highly and moderately resistant, respectively. It would be fairly reasonable to assume that the former two species might also be resistant, because Taiwan is known to be inside the centre of origin of
P. cinnamomi
(6) and they, especially
R. formosanum
, could tolerate the high water content of the growing medium. They would be worthy of particular attention as the cross parent for heat resistant rhododendrons.
|
|
Fig. 2. The survival ratio, vigour and growth of the seedlings in the species of Japanese origin.
For details see Fig. 1. |
2. Species of Japanese origin
The results are shown in Fig. 2. As compared with the foreign species, the characteristic growth pattern of the Japanese species was a projection in the direction to survival and vigour, indicating the rosette type-early growth of seedlings. This tendency was also seen in the species of Taiwan origin.
Concerning heat tolerance there occurred similar variations in the species of Japanese origin to those of foreign origin. As for
R. aureum
, only one form from Kitadake of Yamanashi Prefecture could be tested, but the result was the worst as initially expected from its general lack of heat tolerance. In other species, great intraspecific variation was encountered with the difference of seed sources concerned. If this variation is based on the genetic difference within each species, more detailed and systematic surveys throughout Japan will be needed, including the clarification of the following two issues, namely where the best seed source is and what individual is the most promising as the cross parent for future breeding.
As far as the present experiment is concerned, however,
R. metternichii
var.
hondoense
f.
brevifolium
was the most heat tolerant Japanese species. This form is an inhabitant of Oki-islands, the small island of Shimane Prefecture located at the southern part of the Sea of Japan, where it grows from almost sea level to the mountain tops, the highest of which is only 608 m.
This geographical isolation without high mountains might afford heat tolerance to this form, as compared with the typical
R. metternichii
and
R. yakushimanum
which have high mountains to climb (1700 m or over). Furthermore, some of the finest individuals of this form show compact growth with very shiny, shorter leaves that are elegantly curved down at the edges. One of the authors (K.A.) rates this form rather higher than
R. yakushimanum
combining heat resistance with excellent foliage and flowers.

|
|
Fig. 3-1. The survival ratio, vigour and growth of the seedlings in the hybrids including R.
yakushimanum within their pedigree. For details see Fig. 1. |
3. Hybrids
As in the case of species rhododendrons, the hybrids also showed great diversity in their heat tolerance in accordance with the difference of cross-combinations (Figs. 3-1 and 3-2). Dostãlek etal. (2) had found similar variations especially in vigour and uniformity, in the hybrid seedlings obtained from the diallele cross and self-pollination of a few species and a cultivar ('Cunningham's White'), although the main purpose of their investigation was to disclose the combining ability to produce F1 hybrid seed of rhododendron rootstocks.
It was difficult, however, to draw general conclusions from the present data obtained, mainly because, as expected from the seed sources, each cross combination was not so elaborately planned to compare each other systematically, and the individual used for the cross parent was not necessarily the same individual in such species as
R. yakushimanum
,
R. degronianum
and
R. hyperythrum
.
In some cross combinations, however, consistent results were obtained; the hybrids derived from
R. hyperythrum
and those from 'Scandinavia' gave consistently good results, whereas all of those from 'Crest' were rather poor. This might suggest that some specific species or cultivars inherently have high heritability for heat tolerance and the others have not.
As in the other countries, the special emphasis on
R. yakushimanum
and its derivative hybrids to be used as the cross parent, prevails in this country, chiefly because of their excellent ornamental values (Fig. 3-1). In this country, however, it is rather desirable that this emphasis should be turned to
R. hyperythrum
, as far as the heat tolerance is concerned.
In the other combinations the followings gave good results;
R. yakushimanum
x 'Elizabeth',
R. yakushimanum
x 'Ruby Hart', 'Mrs. Furnival' x
R. yakushimanum
, 'Scintillation' x
R. yakushimanum
, 'Venus' x
R. yakushimanum
, ('Britannia' x
R. yakushimanum
) x
R. yakushimanum
, (
R. yakushimanum
x 'Vulcan') x 'Jock',
R. degronianum
x 'Bow Bells' and
R. metternichii
var.
hondoense
f.
brevifolium
x 'Gill's Crimson'. The definite reason why these combinations gave good results is not clear. As mentioned before,
R. metternichii
var.
hondoense
f.
brevifolium
showed good performance. The descendants of this form also gave consistently good results in several other combinations than that of 'Gill's Crimson', although the detailed data is not presented here. Therefore, this form appears to provide promising material for heat resistant rhododendrons.

|
|
Fig. 3-2. The survival ratio, vigour and growth of the seedlings in the hybrids not including R.
yakushimanum within their pedigree. For details see Fig. 1. |
Summary
The present investigation was conducted to get some clues to breeding for heat resistant rhododendrons from the comparisons of the growth-differences in the seedlings of various species and hybrids. In both species and hybrids, the great diversity in the seedling growth was encountered with the difference of series, subseries, species, varieties, seed sources or cross-combinations.
A large number of species were revealed to be inappropriate to be used as the cross parent for future breeding. Some species, however, seemed to be promising on the basis of the performance of their own and, in some cases, together with their descendants. They were
R. formosanum
,
R. hyperythrum
,
R. metternichii
var.
hondoense
f.
brevifolium
and
R. ponticum
and, to a lesser extent,
R. decorum
,
R. diaprepes
,
R. discolor
,
R. elliottii
,
R. fortunei
and
R. morii
.
In some species, the possibility was suggested that heat tolerance had been secured by the geographical dispersal of long distances from the centre of origin and/or the isolation into small islands without high mountains to climb. It was also suggested that some specific species or cultivars inherently have high heritability for heat tolerance and the others have not.
References
1. Arisumi, K., Matsuo, E. and Sakala, Y.: Breeding for the heat resistant rhododendrons.
Mem. Fac. Agr. Kagoshima Univ.
15, 73-78 (1979)
2. Dostãlek, J. and Dostãlokovã, A.: Unterlagen für Rhododendron aus Hybridsaatgut.
Z. Pflanzenzüchtg.
67, 120-130 (1972)
3. Hoitink, H.A.J. and Schmitthenner, A.F.: Relative prevalence and virulence of Phytophthora species involved in rhododendron root rot.
Phytopathology
64, 1371-1374 (1974)
4. Hoitink, H.A.J. and Schmitthenner, A.F.: Resistance of Rhododendron species and hybrids to Phytophthora root rot.
Bull. Amer. Rhod. Soc.
29, 37-41 (1975)
5. Kanehira, R.: Formosan trees, p. 538-539, Inoue Book Co., Tokyo (1936)
6. Ko, W.H., Chang, H.S. and Su, H.J.: Isolates of
Phytophthora cinnamomi
from Taiwan as evidence for an Asian origin of the species.
Trans. Br. Mycol. Soc.
71, 496-499 (1978)