Introduction
Upper leaf surface (adaxial) hairs on unfolding leaves of rhododendrons in the subgenus Hymenanthes
have captured my botanical curiosity lately.
My first encounter with leaf hairs was through reading Contributions Toward a Classification of Rhododendron
in the Proceedings of the International Rhododendron Conference, sponsored by the New York Botanical Garden and
the American Rhododendron Society in May 1978 (Luteyn J.L. and Mary E. O'Brien, Editors. 1980). Hair types have
been studied using the light microscope by many botanists such as Cowan (1950) and by Chamberlain, Cullen, Hedegaard,
Philipson, Sleumer, Seithe, von Hoff and others who clearly established the presence of several hair types on many
plant parts in different subgenera of genus Rhododendron. The most striking differences in hair structure
are found on the under (abaxial) leaf surface; the hair types on the under leaf surfaces can be taken as being
characteristic for Hymenanthes species (Seithe, et al., 1978).
The classification of Rhododendron follows Sleumer's and Chamberlain's system (Chamberlain et al 1996).
This predominantly morphologically trichome based system had been widely accepted by many in the rhododendron
world (Helm, 2000). Recent DNA research is changing the morphological system of rhododendron classification,
but trichome morphology is retained in the classification of subgenus Hymenanthes and subgenus
Rhododendron. (benhall@u.washington.edu 2006).
Later on, I viewed the late Art Dome's photographs of "those incredible hair-like epidermal appendages" in his
four-part series in the Journal American Rhododendron Society in 1999-2000. Dome's photographs inspired me
to attempt to explore the miniature world of the incredible leaf appendages. Thirteen of Dome's Hymenanthes
photos show dense indumentum on the under (abaxial) leaf surfaces. One of his photos shows stalked glandular hairs
on an upper (adaxial) leaf surface that disappear as the leaf matures. Two photos of two different species show
branched hairs on the upper leaf surface that also disappear as the leaf matures.
Indumentum is defined as "a wooly or hairy covering of leaves or young shoots." It can be on both top (adaxial)
and bottom (abaxial) surfaces of new or young leaves but is usually confined to the underside of mature leaves.
Indumentum referred to in this article is the densely and usually persistent hair covering on the under (abaxial)
leaf surface of rhododendron leaves, as illustrated for R. degronianum ssp. yakushimanum (Fig. 1).
An under stratum of reduced hair forms such as rosette hairs (hairs that radiate outwards from one point) often
occur beneath the larger sized branched hairs of the indumentum, resulting in a two-layered densely packed
indumentum hair cover (Seithe, et al., 1978)
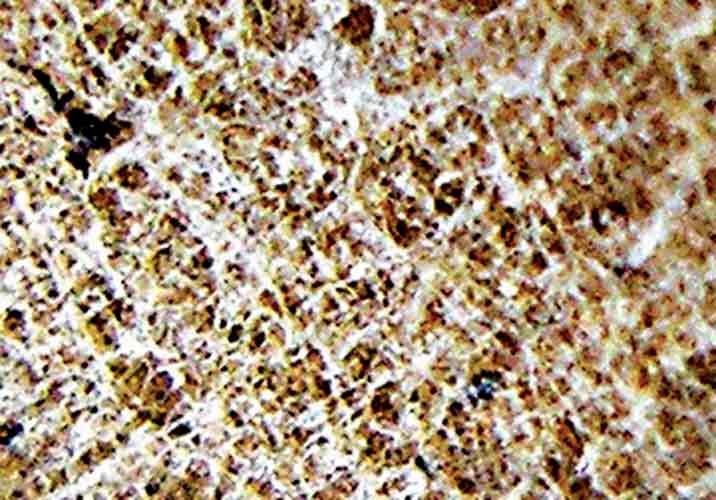 |
| Figure 1. R. degronianum ssp.
yakushimanum with dense under surface branched hairs. Photo by Willem Morsink |
Photographs of exfoliating hairs on upper leaf surfaces are few in number. It is these hairs that can easily be photographed from unfolding leaves. Hairs on R. 'Calsap' (Fig 2) with branched hairs, and Birkett's unnamed hybrid (Fig 3) with stalked globular glands lead a fleeting existence and later disappear (exfoliate) to produce mostly glabrous upper leaf surfaces at maturity on several rhododendrons. The upper leaf surface of R. 'Gustav Melquist' (Fig. 4) shows rusty brown branched hairs that exfoliate in autumn. R. 'Schneekrone' (Figs. 5 and 6) which features under (abaxial) leaf surface indumentum maintains hair cover on the upper (adaxial) surface till later in the year when hairs exfoliate. Two-year-old glabrous leaves (Fig. 5) show a deep green colour on the upper leaf surface.
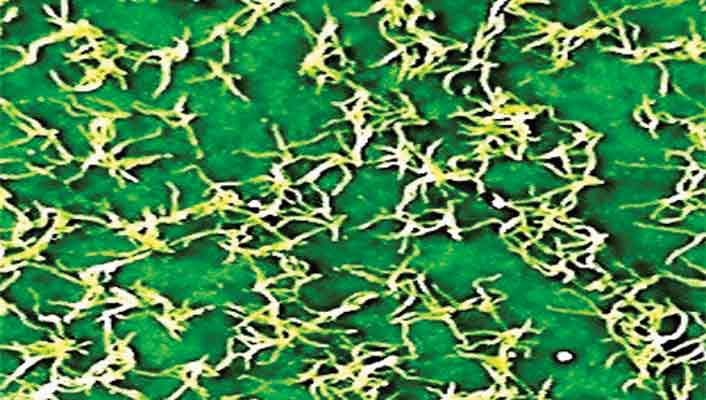 |
| Figure 2. 'Calsap' with upper leaf
surface branched hairs. Photo by Willem Morsink |
 |
| Figure 3. Unnamed hybrid by Richard
Birkett with stalked glands. Photo by Willem Morsink |
Immature leaves of many if not all Hymenanthes rhododendrons have already pre-formed hairs within leaf
buds (Seithe and von Hoff, 1978). These preformed hairs become visible as the immature leaf unfolds. My focus
and point of interest became to explore this fleeting and temporary presence of hairs on upper leaf surfaces
of Hymenanthes rhododendrons. But viewing these minute hairs is a challenge. Ordinary 10-X hand lenses
will show the felt-like indumentum produced by hairs, and the scaly nature of lepidote rhododendron leaves,
but this low magnification is inadequate to show individual hair form. I pondered how I would be able to see
hair form without the use of a light microscope as used by several researchers (Luteyn and O'Brien, 1980).
Hairs growing on the upper surface of leaves are not nearly as densely packed as the under-leaf surface.
Dome's photographs showed that upper leaf surface hairs could be photographed without the use of a light
microscope. I therefore looked at the digital camera for a solution. An Olympus 5.1 mega pixel camera with
an essential super macro mode for close-ups (as close as 2 cm from above the leaf surface) and with pictures
down loaded to a Windows-2000XP computer and using the "Picture It" program solved the dilemma on how to
view the minute epidermal appendages. This method is presented in this paper.
 |
| Figure 4. 'Gustav Mehlquist' with upper
leaf surface exfoliating brown hairs. Photo by Willem Morsink |
 |
| Figure 5. 'Schneekrone' with no surface
hairs remaining on last year's leaves. Photo by Willem Morsink |
 |
| Figure 6. 'Schneekrone' with upper leaf
surface and exfoliating white hair cover. Photo by Willem Morsink |
Hairs: A Brief Summary
Hairs are also known as trichomes; they are found on numerous flowering plant families (Esau, 1977). Trichomes
can be unicellular, multicellular, and glandular or they occur as peltate scale hairs. Other types of appendages
occur in many other plant groups. Hairs or trichomes occur on all plant parts and may persist or fall off. Some
of the persistent hairs remain alive, others die and become dry. Esau, in her review of trichomes (hairs) on seed
plants, notes that they are highly variable structures but also that trichomes can be remarkably uniform in a given
taxon, which is useful for classification.
All rhododendron trichome types begin their development as epidermal cells (Seithe and von Hoff, 1978). In subgenus
Rhododendron the lepidote scale hairs turn their cell axes horizontal to the leaf surface in subsequent
cell growth resulting in a broad flat structure on a short pedestal, the scales.
In subgenus Hymenanthes, branched hairs (Fig. 2) and gland hairs (Fig. 3) hairs occur. Branched hairs
turn their axes perpendicular to the leaf surface. Seithe observed that species that do not have glandular hairs
on mature leaves often have them on the margins of the cotyledons (the two seed leaves in dicotyledonous plants)
and the epicotyl (upper portion of seedlings above the cotyledons ). Erik Nilsen, Ph.D., Professor of Biology at
Virginia Tech, Blacksburg, VA, published numerous papers on a number of different rhododendron topics; he researches
trichomes and published an interesting research article on trichomes of Vireya rhododendrons in the ARS Journal.
In this study it was noted that the significance of scale hairs typical of subgenus Rhododendron and their
relationship with leaf stomata on lower (abaxial) leaf surfaces had not been established by previous research (Nilsen
and Webb, 2007). They explored and investigated the question: Does scale type and density of Rhododendron
species of section Vireya have any relationship to the transpiration function of stomata? Their study
indicates that further work on such relationships is needed before a theory about scale influence on stomatal function
can be finalized.
Possible significances of trichomes include reducing vapor demand by increasing humidity in the air above stomata,
reducing palatability of leaf tissue to arthropod herbivores and increasing leaf reflectance of light from the upper
(adaxial) leaf surface. This research paper has an interesting introduction on plants solving the dilemma of
balancing water loss during carbon (CO2) uptake and how stomata and possibly trichomes may be involved.
There exist many works on trichomes and an elaboration of this topic is better left to scientists "in the know." My
focus will be limited to a visual digital photographic exploration of upper-leaf surface (adaxial) hairs on unfolding
Hymenanthes rhododendrons.
Digital Photography
There are many different makes of digital cameras, and the better ones usually feature a "close-up" feature which is
essential for hair photography. I use an Olympus 5.1 mega pixel camera attached to a mini-tripod and photograph leaf
surfaces at a distance from 2 to 2.5 cm, and download pictures to the Windows-2000 XP computer with a "Picture It"
program. Obtaining successful close-ups of leaf appendages (hairs) requires a number of steps in preparing the
Olympus camera for action.
1. Use the automatic setting on the mode dial to take your picture; experienced photographers can use the manual
method described in advanced manuals. With the camera battery power switched on, press the Ok-Menu button inside
the Arrow Pad and select the left arrow lever showing SQ2, SQ1, HQ, and SHQ choices. Select SHQ for the highest
Pixel level of 2592 x 1944 pixels and click Ok-Menu button.
2. It is essential to switch on the super macro mode (indicated with S and tulip symbol), for close-ups of hairs
on leaf surfaces measuring from 2.5 x 3.4cm and as close as 2 cm from above the leaf surface. Use the bottom
"direct" button located in a row of four buttons to the left of the arrow pad.
3. Using the viewing monitor place the target marker on the leaf area of interest. To automatically focus on the
subject press the shutter button halfway down and hold; a green light on the screen comes on when the focus and
exposure are locked. Next press the shutter button fully down and an orange light beside the view finder blinks
while the picture is recorded on to the chip card.
4. Down load your pictures to the Window XP PC, using a picture download cable. The Windows should have a Wizard
download program. The program allows you to load the pictures into a folder to be stored in your Pictures file.
5. The Windows has a "Picture It" program which I use; (Olympus also provides its own picture edit program). Using
the Edit Picture program I select a leaf from my hair-picture folder and zoom it to a desired magnification up from
20 to 100% or more. I crop the pictures to a standard wallet size of 3 x 5, sharpen the image and contrast and
then save it for later use as illustrated in Figs. 7, 8.
 |
| Figure 7. 'Paul R. Bosley'
with leaf at low magnification Photo by Willem Morsink |
 |
| Figure 8. 'Paul R. Bosley' with selected
leaf area magnified Photo by Willem Morsink |
Leaf Branched Hairs
I found that many of the so-called "glabrous-leaved plants" that I examined feature (adaxial) upper surface
exfoliating hairs - hairs that flake-off sooner or later in the same year. New unfolding leaves show simple
branched hairs for R. 'Wyandanch Pink' (Fig. 9) and a seedling of R. maximum 'Leach's Mount Mitchell'
(Fig. 11). Branched hairs occur only sparingly on R. 'Cunningham's White' and R. 'Scintillation'.
Plants such as 'Saint Michel', which features persistent under leaf surface indumentum, has pre-formed hairs
on the upper surfaces of unfolding leaves. These upper-surface hairs remain much longer (until late fall), as
compared to plants without under-leaf surface indumentum.
Leaf-Gland Hairs
A glandular hair has a small spherical knob of sticky secreting tissue on the top of the hair (stalk).
Glandular hairs are found on Hymenanthes plants and have a long or short stalk that bears a
multi-cellular orange to brown-tipped gland (R. 'Paul R. Bosley' Fig. 10), a red-tipped gland (R. 'Ponticum
Roseum'), and a white-tipped gland (R. 'Nova Zembla'). The white coloured glands possibly indicate
desiccated tissue.
Stalked glands may occur together with the branched hairs for the upper leaf surface of R. maximum
'Leach's Mount Mitchell' and R. smirnowii, or less frequently only stalked glands grow on the
upper leaf surface (Fig. 3).
Gland-like hairs on R. 'Helsinki University' can be seen growing from branched hairs on the upper leaf
surface. These gland hairs appear as an open cup with an undulating margin. The branched and/or
glandular-like hairs are present on upper and under leaf surfaces of most of my plants These hairs
mostly disappeared - exfoliated - during the summer.
 |
| Figure 9. 'Wyandanch Pink' with upper
leaf surface with branched hairs. Photo by Willem Morsink |
 |
| Figure 10. 'Paul R. Bosley' with upper
leaf surface of red-brown tipped glands - primitive branched hairs. Photo by Willem Morsink |
 |
| Figure 11. A seedling of
R. maximum 'Leach's Mount Mitchell' with upper leaf surface hairs. Photo by Willem Morsink |
Colour Variation for Gland Hairs of R. maximum from Different Sources:
They show either yellow tipped or red-tipped gland hairs as follows:
Yellow-brown-tipped glands of R. maximum seedling. This plant with white flowers
supposedly is a seedling of red maximum selected by Leach (1964). I obtained a specimen in
1996 as a plant grown from the ARS seed exchange. The plant is reported to have red flower buds
opening to become bright pink flowers, red on the outside with yellow green dorsal blotch (Greer,
1996). My plant presently is about 3 feet tall and starts with faint pink flowers bud changing to
snow white with a greenish blotch and flowers late June and the first week of July in Toronto. The
upper leaf surface shows yellow-brown-tipped glands (Fig. 11) between primitively branched hairs
for spring 2006 when this plant produced flowers. In 2007, there were no flowers and the leaves
show few if any glands but instead many branching hairs. Hairs disappear by the end of July.
Yellow-brown-tipped glands of R. maximum seedling. Primitive branched hairs and yellow-tipped gland hairs are visible on the upper and under leaf surfaces of seedling. This plant is about 6 inches tall, and was grown from an R. maximum seed capsule - seed obtained in the Norfolk Arboretum about 1996.
Red tipped glands of R. maximum seedling. This plant is about 6 inches tall and was grown from a seed of a second R. maximum seed capsule obtained in the Norfolk Arboretum. It shows red tipped glands between a dense cover of white branched hairs on the upper leaf surface of young unfolding leaves as well on the lower leaf surface. Hairs disappear by the end of the summer. Lower leaf surfaces remain a rusty brown colour. This seedling resembles the Tunnel Mountain source with red tipped glandular hairs.
Red tipped glands of R. maximum (Tunnel Mountain). This is a rescued, mowed-over seedling from a picnic roadside area at Tunnel Mountain, Interstate 76-Virginia/North Carolina area. This plant is also 3 feet tall and produces unattractive white flowers during the last week of June into the first week of July. The upper surface has red gland-like appendages. Of interest is that Parker Little (2002) observed a R. maximum on White Rock Mountain, West Virginia, and noted that "one shrub had young leaves with red-purple glands raised above the dense white scurfy layer."
Red tipped glands of 'Roseum', a hybrid between R. ponticum x R. maximum. My plant was purchased about 1999 and was grown by the Looyes, ARS District 12 members in Niagara-on-the-Lake, Ontario. It is vigorous grower and about 3 feet tall. It flowers mid-June in Toronto. I have several additional plants which flower most years. The upper leaf surface from my first plant has branched hairs and bright red tipped gland hairs in addition to a few orange-brown gland hairs elsewhere on the leaf. Is it possible that the red colour of the gland hairs is a genetic contribution from the maximum pollen parent? The varying gland colours of the seedling leaves of R. maximum suggest genetic variability by either the presence of yellow-brown or bright red coloured gland hairs. R. maximum ranges through the Appalachian Smokey Mountain area to southern Maine (www.planthardiness.gc.ca).
Conclusion
Digital photography made it possible to view the infrequently seen world of minute appendages that
occur on the surfaces of rhododendron leaves for a short leaf unfolding period early each spring.
My Olympus digital camera with a "Close-up (2cm)" feature in combination with my Windows XP computer
allowed me to obtain a much improved view of hairs on my rhododendrons and to initiate my hair
exploration.
During my June and July 2007 visit to British Columbia I searched for rhododendrons unfamiliar to
eastern North Americans. It was too late for R. macrophyllum, which finished flowering
by early July, but its seedlings from the same area showed hairs on the cotyledons. R.
hemsleyanum was in full flower when I visited in late June, but the new shoots were only in
early stage of development and had not yet differentiated its future leaves. Previous year's
leaves were glabrous on this species. Hairiness of upper leaf surfaces of this and other glabrous
species of the temperate North American West Coast remains to be determined in future years.
Hairs on the upper (adaxial) surfaces of so-called "glabrous-hairless" species may have been
under reported because of the taxonomist's focus on hair-indumentum on under (abaxial) leaf
surfaces in classification. Could exfoliating hairs that flake-off after a short time period be
utilized in classification, or in distinguishing between hybrids and cultivars and, if not,
what could their function be as they appear persistently on numerous Hymenanthes plants?
Upper surface hairs were fleetingly visible on scores of Hymenanthes rhododendrons in this
exploration. The hairs then proceeded to slough off during the same growing season and do not
usually appear to be transformed for one or another function. One trichome function had been
indicated to possibly be involved with reducing vapor demand by increasing humidity in the air
in the vicinity of stomata, the small leaf surface openings on the lower side of leaves;
rhododendrons predominantly feature stomata on the lower leaf surfaces (Nilsen and Webb,
2007). As there are no upper surface stomata on Hymenanthes rhododendrons, the
leaf-hairs are not needed for this function and presumably disappear for this reason. Yet,
hairs both for upper and under leaf surfaces continue to be preformed within buds. The
question remains why do rhododendrons persist in pre-forming hairs (trichomes) on the
miniature leaves within buds? My speculation as a lay person is that gland hairs could be
relic-non-functioning reproductive structures of ancestral rhododendrons, before the
calyx, corolla-stamen, and ovary-carpel floral solution differentiated from floral
meristems.
References
Chamberlain, David, Roger Hyam, George Argent, Gillian Fairweather and Kerry S. Walter, 1996.
The Genus Rhododendron, Its classification & synonymy. Edinburgh: Royal Botanic Garden,
Edinburgh.
Cowan, John Macqueen. 1950. The Rhododendron Leaf. Edinburgh: Oliver and Boyd.
Dome, Arthur. 1999. Subgenus Hymenanthes Part I, Jour. Amer. Rhod. Soc. Vol. 53(4): 198-9.
__________ 2000. Subgenus Rhododendron, Part II, Jour. Amer. Rhod. Soc. Vol.54 (1): 42-3.
__________ 2000. Subgenus Hymenanthes Part III, Jour. Amer. Rhod. Soc. Vol.54 (2):78-9.
__________ 2000. Subgenus Rhododendron Part IV, Jour. Amer. Rhod. Soc. Vol.54 (3):138-9.
Esau, Katherine. 1977. The Anatomy of Seed Plants. New York: John Wiley& Sons.
Greer, H.E. 1996. Greer's Guidebook to Available Rhododendrons-Species and Hybrids. 3rd Edition.
Eugene, Oregon: Offshoot Publications.
Hall, Ben. 2006. Research Review-Rhododendron taxonomy. Jour. Amer. Rhod. Soc. Vol.60 (3):124
Helm, Henry R. 2000. Rhododendrons? Azaleas? Why and where have all the azaleas gone? Jour. Amer.
Rhod. Soc. Vol.54 (1):9-10.
Little, Parker L., 2002. An exploration and study of White Rock Mountain, WV for unusual forms
of Rhododendron species. Jour. Amer. Rhod. Soc. Vol. 56 (3): 122-131
Luteyn J.L. and Mary E. O'Brien, Editors. 1980. Contributions toward a Classification of
Rhododendron. Proceedings of the International Rhododendron Conference. The New York Botanical
Garden, May 15-16, 1978. Sponsored by the New York Botanical Garden and the American
Rhododendron Society
Nilsen, Erik T. and David Webb. 2007. Does scale type and density of Rhododendron species of
section Vireya have any relationship to stomata? Jour. Amer. Rhod. Soc. Vol. 61(2) 93-103.
Seithe, Almut and Nee von Hoff. 1978. In: Luteyn, J.L. and Mary E. O'Brien, Editors, 1980.
Proceedings of the International Rhododendron Conference. The New York Botanical Garden,
May 15-16, 1978. Sponsored by the New York Botanical Garden and the American Rhododendron Society.
Willem (Bill) Morsink is a Toronto member of ARS District 12.