
QBARS - v35n1 Physical and Chemical Responses of Naturally Grown Rhododendron to Environmental Stresses
Physical and Chemical Responses of Naturally
Grown Rhododendron to Environmental Stresses
H. L. Gholz
School of Forest Resources and Conservation
University of Florida Gainesville, FL
INTRODUCTION
The western rhododendron (
Rhododendron macrophyllum
G. Don) is one of the most cosmopolitan shrub species of the coniferous forests in the western Cascade Mountains of Oregon. Rhododendron occurs naturally at all elevations, in all but the coldest and hottest communities, and is the most important under story species in many of the very old (400+ years) hemlock and Douglas-fir stands. It is also abundant on clear cuttings, where it re-sprouts and persists for years until it again is in the shade.
Rhododendron's success in such a wide range of changing microclimates is obviously due to a great plasticity in growth form and reproduction. It is therefore an ideal study object for examining how different levels of environmental stress effect the appearance and growth of a plant species in natural and disturbed areas.
STUDY AREAS
Rhododendron in the under story of three undisturbed old-growth forests (at low, middle, and high elevations), dominated by 450-year-old Douglas-fir (
Pseudotsuga menziesii
Mirb. Franco), and in one 1 5-year-old clear-cutting were chosen for study. All four sites were located within the H. J. Andrews Experimental Forest (44° N, 122° W), and represented typical coniferous communities. There were no major soil chemical differences among the sites chosen for this study, and at all four locations rhododendron was the most abundant shrub species.
A clear-cutting (plot 77) was selected to compare the effects of full sun on this species, which occurs naturally in heavily shaded environments. The area was logged in 1960, and the rhododendron re-sprouted heavily the following year from preexisting root masses.
The three under story communities selected composed a temperature and growing-season gradient from a low-elevation, warm site with the longest growing season (plot 17), through a cool, mid elevation site (plot 5), to a cold, upper elevation site with the shortest growing. season (plot 12). The shrub layers in all three sites experienced light intensities during the growing season of 3 to 7% full sun (clear-cutting = 100%).
Also, a transect of plots increasing in light exposure at the shrub surface was established from within the under story of plot 5 into an adjacent clear-cutting. Light levels measured in late June on these plots ranged from 1 to 100% full sun. This light transect furnished samples so that the effects of increased light on leaf variables could be determined while other factors such as soil type, slope-and aspect, elevation, and precipitation remained constant.
RESULTS
There were no strong trends at any of the sites in percent or per surface area nitrogen (N) or phosphorus (P) concentrations with season other than a decrease in current foliage. However, there were large differences among the locations, especially for physical characteristics of the leaves.
|
|---|
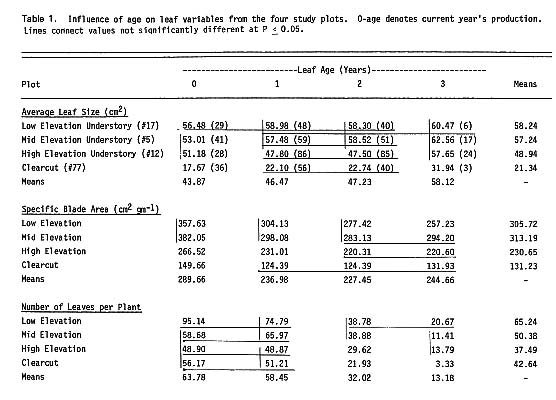
Table 1 summarizes the differences in leaf characteristics related to light, elevation (which affects growing season length and temperature), and leaf age. The values represent the seasonal means for each plot and leaf age class. Although there were few leaves more than 3 years old on the clear-cut plants, plants of similar diameter had a greater total number of leaves than plants in the high-elevation under story, emphasizing the importance of leaf size and its sensitivity to increased light levels. At all locations, average leaf size increased with leaf age. The pattern was reversed for specific leaf area (cm 2 g -1 ) which decreased with age except on the clear-cutting where it reached a plateau after 1 year. Average leaf size and specific area both decreased markedly at the high-elevation and clear-cut plots.
For an average plant (1.5 - 2.0 cm basal diameter) on the four plots, the leaf area varied by a factor of 3, while foliage biomass (total foliage tissue) for the same plants varied only by a factor of 1.8. There were no significant differences in height as a function of basal diameter, except that low-elevation under story (17) plants were taller for any given diameter than plants in the other three areas, and no differences were found among sites for woody biomass (total woody tissue) per stem. However, 15-year-otd plants on the clear-cutting had woody biomass equal to shaded plants from 25 to 60 years old, which indicates either that stem growth rates are two to four times as great on the clear-cutting or that the plants will show an abrupt decrease in growth rate with age, a pattern not detectable at 15 years.
|
|---|
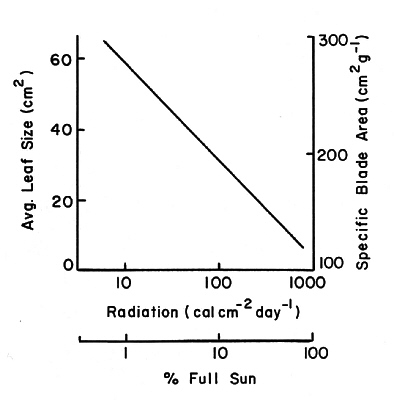
Fig. 1. Pattern of leaf dimensions from plants 1.5 - 2.0 cm basal diameter sampled along the light transect from the under- story of plot 5 into an adjacent clear cutting. |
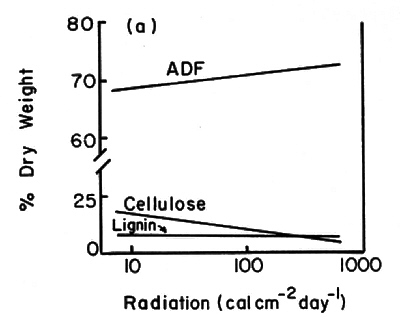
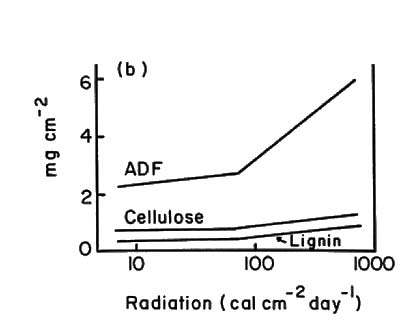
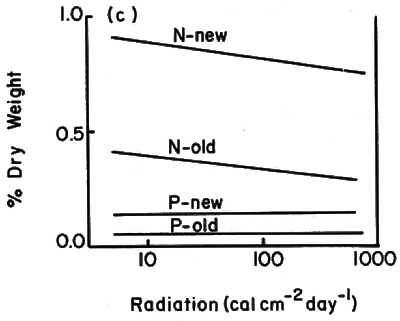
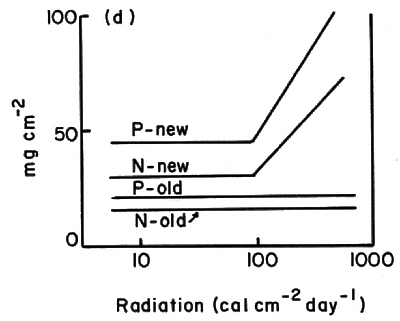
Along the light transect average leaf size and specific area decreased by factors of four and three, respectively, from 1% of full sun to full sun (Fig. 1). Soluble carbohydrates (acid detergent fraction or ADF), and nitrogen and phosphorus in current foliage showed large per unit-area increases at >10% full sun (Fig. 2b and d). Percent concentrations showed little change over this gradient (Fig. 2a and c).
|
||||||
|---|---|---|---|---|---|---|
Fig. 2. Patterns of leaf fiber content, cellulose and lignin percent dry weights (a) and mg per cm 2 (b); and, patterns of N and P contents in new and old foliage percent dry weight (c) and mg per cm? (d). All samples were from plants 1.5 - 2.0 cm basal diameter sampled along the light transect at plot 5. |
DISCUSSION
|
|---|
Several characteristics of the clear-cut, or sun, rhododendron illustrate adaptations to the exposed site. In order to survive, they have restricted their photosynthesizing-transpiring surface area. The total leaf area per plant on the clearcutting averaged less than half that of any of the shaded plants (Table 2). Leaf size was even smaller than on the high-elevation shaded site (Table 1). Percentage foliage concentrations of nitrogen and phosphorous on the clear-cut (77) plants were statistically less than on under story (17) plants (rarely more than 0.75% N or 0.07% P in other than current foliage). In addition some indirect evidence supports the idea that rhododendron is inefficient in its water uptake and conducting system in comparison with some of its associates (blaring et al., 1977).
Cooler temperatures, a shorter growing season, and a deeper and longer lasting snow pack at the high-elevation under story site (12) may limit total leaf area (and to lesser extent, the leaf biomass) of rhododendron plants growing there. This limitation could be a result of selective pressures to reduce snow damage by lowering resistance to the snow pack. Both smaller leaves and fewer leaves helped reduce the leaf area and biomass at the highest elevation under story site (12).
Expression of chemical concentrations on a unit area may be a valuable alternative to conventional expression as percentage dry weight and may more realistically reflect physiological stresses. 'Optimal' percentage dry weight levels of foliar nutrient concentrations have rarely been demonstrated satisfactorily under natural conditions, mainly because of the large variation observed (often because of nonstandard sampling procedures).
In this study, structural features of the leaves and total plant leaf surface areas were the best indicators of stress. Average leaf size, specific leaf area, number of leaves, and plant leaf area all decreased markedly with increasing stress (represented by high elevations and exposure to full sun). Lignin, cellulose and the ADF per cm 2 leaf blade area all increased substantially at light levels
10% of full sun. N and P % concentrations were highly variable. N and P concentrations per cm
2
showed only slight differences with site and leaf age, although current foliage of exposed plants had values 3-4 times higher than current foliage of shaded plants.
LITERATURE CITED
GHOLZ, H. L. 1976. Assessing stress in Rhododendron macrophyllum through an analysis of leaf physical and chemical characteristics. Can. J. Bot. 56:546-556.
WAKING, R. H., H. L. GHOLZ, C. C. GRIER, and M. L. PLUMMER. 1977. Evaluating stem conducting tissue as an estimator of leaf area in four woody angiosperms. Can. J. Bot. 55(1 1):1474-1477.
