JARS v44n4 - Influence of Light and Temperature on Seed Germination of Flame Azalea
Influence of Light and Temperature on Seed Germination of Flame Azalea
Asiah Malek, Frank Blazich, Stuart Warren, and James Shelton
North Carolina State University
Raleigh, North Carolina
Reprinted from the Journal of Environmental Horticulture, Spring 1989
Abstract
Seeds of flame azalea [
Rhododendron calendulaceum
(Michx.) Torr.] were germinated at 25°C (77°F) and 25°/15°C (77°/59°F) with daily photoperiods of 0, ½, ½ twice daily, 1, 2, 4, 8, 12 and 24 hours. Seeds exhibited an obligate light requirement. At 25°C (77°F), increasing photoperiods increased germination with maximum germination (85%) occurring by day 12 under continuous illumination. The alternating temperature of 25°/15°C (77°/59°F) enhanced germination when light was limiting. At this temperature, maximum germination of 84 to 91% was reached by day 24 for all photoperiods
>
½ hour although at photoperiods > 4 hour comparable germination was noted at day 18.
Introduction
Flame azalea [
Rhododendron calendulaceum
(Michx.) Torr.] is indigenous to the Appalachian region of the United States, extending from southwestern Pennsylvania to northern Georgia (5, 6). It blooms in late spring and is regarded as one of the most striking native flowering shrubs, with flower color ranging from orange-yellow to scarlet (6).
Although the commercial potential of this outstanding flowering shrub has long been recognized, it has not been widely utilized due in part to lack of information regarding both sexual (seed) and asexual gs, successful propagation by seed is common. Nurserymen in western North Carolina propagate the species by seeds collected from native plants. However, it appears no quantitative data are available regarding the influence of various environmental factors on germination. Therefore, the objective of this investigation was to examine the influence of light and temperature on seed germination of flame azalea.
Materials and Methods
On November 13, 1986 mature seed capsules were collected from open-pollinated plants growing in Watauga County, North Carolina, at an elevation of 1,400 m (4593 ft). Capsules were stored in a paper bag at 20°C (68°F) for 21 days. Seeds were then removed from the capsules and stored at a moisture content of 6% in a sealed bottle at 4°C (39.2°F). Moisture content of the seeds was determined by calculating the mean moisture content of six 100-seed samples following drying at 105°C (221°F) for 24 hours. In October 1987, seeds were removed from storage and graded by manual removal of abnormal, damaged and undersized seeds. Graded seeds were sown in covered, 9-cm glass Petri dishes containing two pre-washed germination blotters moistened with tap water. Following placement of seeds in the dishes, half were designated for germination at 25°C (77°F) and the other half for germination at an 8/16 hour thermo-period of 25/15°C (77/59°F). All dishes were placed in black sateen cloth bags and the seeds allowed to imbibe overnight prior to initiation of the photoperiod treatments. Dishes were randomized within two growth chambers [C-chambers (4)], set at the appropriate temperatures. Chamber temperatures varied within + 0.5°C (0.9°F) of the set point.
Within each temperature regime, seeds were subjected daily to the following nine photoperiods: total darkness, ½, two ½ hour photoperiods separated by 7½ hours of darkness, 1, 2, 4, 8, 12 and 24 hours. Growth chambers were equipped with cool-white fluorescent lamps which provided a photosynthetic photon flux density (400-700 nm) of 42 µmol m -2 s -1 (3.2 klx) as measured at dish level with a cosine corrected LI-COR LI-185 quantum/radiometer/ photometer. All photoperiod treatments, except the 24 hour illumination, were regulated by removal and placement of the Petri dishes in black sateen cloth bags. For the 24 hour photoperiod treatment, the Petri dishes remained continuously un-bagged in open chamber conditions. Regardless of the photoperiod, temperatures within the Petri dishes as measured with a thermocouple never exceeded ambient by more than 1°C (1.8°F). Petri dishes representing the total darkness treatment were kept in the black cloth bags throughout the experiment and all watering and germination counts were performed under a green safelight. Germination blotters were kept moist with tap water throughout the experiment. Seeds showing signs of decay were immediately removed from the dishes. Each photoperiod treatment was replicated four times with a replication consisting of a Petri dish containing 100 seeds. Germination counts were recorded every 3 days for 30 days. A seed was considered germinated when the emerging radicle was > 1 mm (0.04 in.).
Percent germination was calculated as a mean of 4 replications per treatment. Comparison of means was by LSD, at the 5% level.
|
|
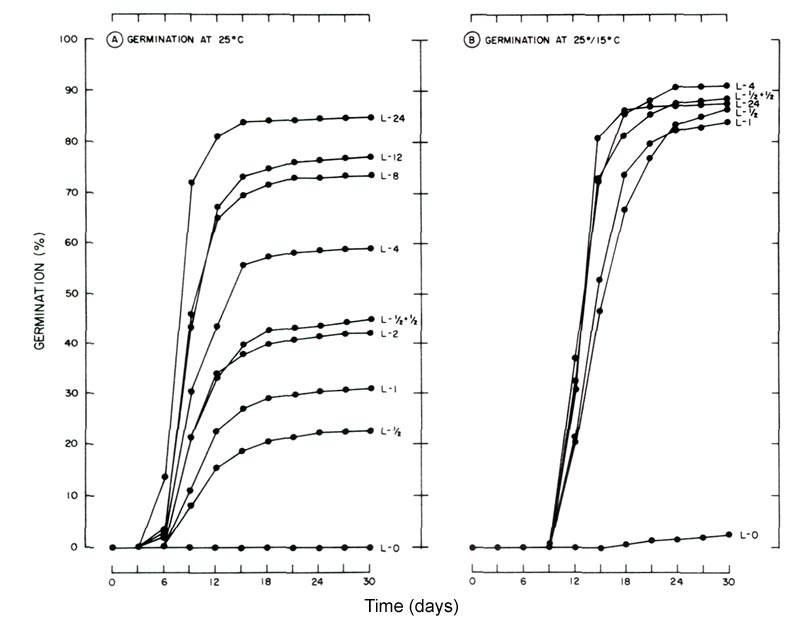
Figure 1. Influence of light and temperature on seed germination of flame azalea. LSD 005 = 3.5
for comparing means within a temperature and between temperatures. (A) germinated at 25°C (77°F) with photoperiods (L) ranging from total darkness (L-0) to 24 hr (L-24). (B) germinated at 25°/15°C (77°/59°F) utilizing the same photoperiods as in A. Data for photoperiods of 2, 8 and 12 hr were omitted since germination was similar to that of the 24-hr light treatment. |
Results and Discussion
Seeds of flame azalea have an obligate light requirement for germination (Fig. 1). This supports previous mention of the need for light to achieve germination of other species of
Rhododendron
(2, 3, 8). However, these reports lack such critical information as the number of hours required daily and the interaction of light and temperature.
At 25°C (77°F), germination commenced earlier than at 25°/15°C (77/59°F) and increasing photoperiods increased germination. By day 30, seeds subjected to the 24 hour photoperiod at 25°C (77°F) attained the highest total germination (85%), which was significantly greater than those receiving the 12 hour photo-period (77%) (Fig. 1A). Maximum germination was reached by day 12 under the 24 hour photoperiod as there was no significant difference in germination from day 12 to 30. There was no significant difference in total germination at the 12 hour photoperiod compared to 8 hour. Seeds receiving daily light treatments < 2 hour never reached 50% germination.
The alternating temperature of 25/ 15°C(77/59°F) enhanced germination when light was limiting (Fig. 1B). For photoperiods > ½ hour germination > 84% was achieved by day 24. However, the earliest point at which comparable germination was reached was at day 18 for photoperiods > 4 hour, day 21 for the ½ + ½, 1 and 2 hour photo-periods and day 24 for the ½ hour photoperiod. For continuous illumination at 25°C (77°F) and for photoperiods > ½ hour at 25°/15°C (77°/59°F), we feel maximum germination was attained because the only seeds remaining in a particular Petri dish at the conclusion of the study were decayed and did not appear to be viable.
The light requirement appeared to be partially overcome by alternating temperature when photoperiod was limiting. In total darkness at 25/15°C (77/59°F), 3% germination was noted at the end of 30 days compared to none at 25°C (77°F). Whether germination would have increased in darkness at 25/15°C (77/59°F) if the experiment had been continued is not known. Also, it is possible germination may have been higher if the differential between the high and low temperature had been greater (2). The relative lack of germination in the dark indicated that the alternating temperature did not fully substitute for the light requirement. It did, however, enhance germination under limiting light levels as observed in several other light-sensitive species (11).
Alternating temperature delayed initial germination by 3 to 6 days compared to the constant temperature. At 25°C (77°F) germination commenced by day 6 for photoperiods > 4 hour and by day 9 for the other light treatments. Initial germination occurred between day 9 and 12 under the alternating temperature.
Under both temperature regimes the twice-daily illumination (two ½ hour exposures separated by 7½ hours of darkness) induced significantly higher germination compared to a single, continuous light treatment of an equivalent photoperiod (Fig.1).
In greenhouse production of seedlings of flame azalea, supplemental light is often used to extend the natural day-length during winter to increase total germination as well as the germination rate (James E. Shelton, personal communication). In the present study, continuous illumination resulted in the highest germination (85%) at 25°C (77°F), although it appears that any photoperiod between 18 and 24 hours would probably have yielded equivalent germination (Fig. 1A).
Based on the results obtained from the alternating temperature treatment, a ½ hour daily photoperiod is sufficient to elicit maximum germination. However, longer photo-periods would undoubtedly be more beneficial for subsequent growth of the seedlings.
|
|
Figure 2. Seeds of flame azalea. Note the variation between
individual seeds. Following drying to a moisture content of 6% there were approximately 122,000 seeds/28 g. (1 oz.). Scale divisions = 0.1 mm (0.004 in). |
Significance to the Nursery Industry
Quantitative data are presented regarding the influence of light and temperature on seed germination of flame azalea. Results show the species requires light for germination. Without daily illumination the seeds will not germinate. Because of the need for light and the small size of the seeds (Fig. 2), nurserymen are advised not to cover the seeds during propagation. Using daily photo-periods equal to or greater than 18 hour maximum germination can be achieved at 25°C (77°F) while comparable results can be realized at 25/15°C (77/59°F) with photoperiods > ½ hour.
Literature Cited
1. Blazich, F.A. and J.R. Acedo. 1988. Micropropagation of flame azalea.
J. Environ. Hort.
6:45-47.
2. Cho, M.S., J.H. Jung, and D.Y. Yearn. 1981. Studies on seed germination of rhododendron plants. J. Korean Soc. Hort. Sci. 22:107-120.
3. Dirr, M.A. and C.W. Heuser, Jr. 1987. The Reference Manual of Woody Plant Propagation: From Seed to Tissue Culture. Varsity Press, Athens, Ga.
4. Downs, R.J. and J.F. Thomas. 1983. Phytotron procedural manual for controlled environment research at the Southeastern Plant Environment Laboratory. N.C. Agr. Res. Serv. Tech. Bul. 244 (Revised)
5. Li, H-L. 1957. Chromosome studies in the azaleas of eastern North America. Amer. J. Bot. 44:8-14.
6. Liberty Hyde Bailey Hortorium. 1976. Hortus Third: A Concise Dictionary of Plants Cultivated in the United States and Canada, 3rd ed. Macmillan Publishing Co., New York.
7. Nolde, S. and J. Coartney. 1985. Clonal variation in rooting of Rhododendron calendulaceum. HortScience 20:539. (Abstr.)
8. Olson, D.F., Jr. 1974. Rhododendron L. rhododendron, p. 709-712. In: C.S. Schopmeyer (Tech. Coordinator). Seeds of Woody Plants in the United States. Agr. Hdbk. 450. U.S. Dept. Agr., Forest Serv., Washington, D.C.
9. Shelton, J.E. and R.E. Bir. 1980. Propagation of flame azalea by softwood cuttings. Proc. Southern Nurserymen's Assoc. Res. Conf., 25th Annu. Rpt. p. 220-221.
10. Skinner, H.T. 1961. Comments on the propagation of native azaleas. Proc. Intern. Plant Prop. Soc. 11:96-98.
11. Toole, E.H., V.K. Toole, H.A. Borthwick, and S.B. Hendricks. 1955. Interaction of temperature and light in germination of seeds. Plant Physiol. 30:473-478.
The authors are associated with the Department of Horticultural Science at the University of North Carolina. A report of Frank Blazich and Juan Acedo's work on the micropropagation of flame azalea appeared in the ARS Journal, Vol. 43:2 (Spring 1989).
