JARS v42n2 - Light Induced Pigmentation Patterning In Rhododendron Corollas
Light Induced Pigmentation Patterning In Rhododendron Corollas
Pat Halligan
Langley, Washington
Did you ever wonder what may be one of the causes of the irregular pattern markings on the petals of the Maddenii hybrids, such as Rhododendron 'Else Frye', the picotee effect of R. racemosum , and the two tone effect of R. glaucophyllum ?
While looking closely at the flowers of the hybrid R. campylogynum x R. spinuliferum , I noticed that the flower was a brick red, but only on the outside of the corolla. Wherever the petals had overlapped in the bud, the flower was white, as was the inside of the corolla. It occurred to me that the color pattern could easily have been caused by light. Wherever light struck the flower in the bud or while it was expanding, the pigment would be activated and cause the flower to turn red. Wherever the flower was shaded, the pigment would not be activated and those parts of the flower would remain white.
Testing the Hypothesis
To test this hypothesis, I placed aluminum foil over buds and flowers of
R. campylogynum
x
R. spinuliferum
, R. 'Mary Fleming' x
R. spinuliferum
,
R. racemosum
,
R. campylogynum
,
R. glaucophyllum
,
R. chameunum
and R. 'Elizabeth'. Depending upon the stage when it was put on, the foil served to exclude light from the developing bud or expanding truss.
Foiled buds of R. 'Mary Fleming' x R. spinuliferum which normally develop into pinkish orange flowers ended up yellowish in color. Foiled buds of R. racemosum and R. glaucophyllum which normally develop into pink flowers ended up almost white (Fig. 1). Placing foil over buds which had already broken open had little or no effect on eventual flower color. Apparently, pigmentation response to light occurs in the expanding bud before the bud breaks open to reveal the flowers.

|
|
Figure 1. Effect of shading with foil on flower color of
R. racemosum . Pink spray has normal unshaded flowers. White spray, from same plant, was shaded by foil, which was left on the branch from the time of bud expansion to opening of flowers. Photo by Pat Halligan |
Placing foil over buds of R. chameunum , R. campylogynum , and R. 'Elizabeth' had virtually no effect on flower color, with the former two flowers becoming a rather uniform purple, and the latter becoming an intense red.
Placing foil over buds of various clones of R. campylogynum x R. spinuliferum resulted in the entire range of effects, depending upon the clone. Of the nine clones tested, four showed a great effect due to shading, two showed a slight effect, and three showed little effect. Those which were greatly affected displayed a characteristic pattern of pigmentation (Fig. 2). The outside of the corolla was reddish except where the petals overlapped in the bud. The inside of the corolla was white. When shaded the flowers ended up white (Fig. 3). Other clones were not affected by shading with foil. These had uniformly pink or red corollas, inside and out (Fig. 4).

|
|
Figure 2. Truss of
R. campylogynum
x
R. spinuliferum
showing a typical light dependent pigmentation pattern. Note the white in the areas of petal overlap and the white visible within the corolla of the flower to the left. Photo by Pat Halligan |

|
|
Figure 3. Truss of
R. campylogynum
x
R. spinuliferum
which
had been shaded from light. The unshaded trusses of this plant showed pigmentation similar to that shown in Figure 2. Photo by Pat Halligan |

|
|
Figure 4. Truss of
R. campylogynum
x
R. spinuliferum
, a
sibling of the plant in Figure 2, showing a light independent pattern. Note that the areas of petal overlap are the same color as the rest of the corolla. The inside of the corolla is similar in color to the outside. Photo by Pat Halligan |
Maddenii Color Patterning
Next, I turned my attentions to the Maddeniis. These plants have large buds which proved to be ideal experimental subjects. Dissected, the buds show certain characteristics which are important in understanding the color patterning in Maddenii flowers (Fig. 5).
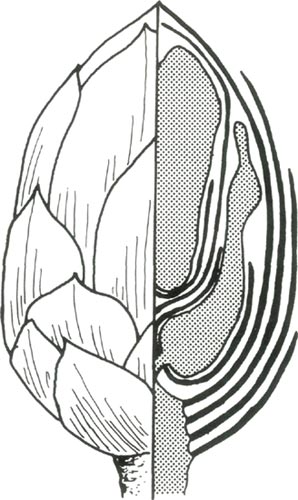
The outer leaf-like coverings are called scales. Although the scales are imbricated (wrapped around) so that they look like shingles, it is important to note that each scale extends all the way from the peduncle, or base of the truss. The effect of this overlapping is that the bud is covered by many layers of scales at the base, but the covering of the bud gets progressively thinner toward the apex. Thus presumably more light penetrates the bud in the apical region than in the basal region. In addition, the calyx is relatively large in the bud and further shades the basal area of each flower. Between the flowers are bracts that also have some effect upon shading within the bud.
|
|
Figure 5. Diagram of dissected bud of
R. nuttallii
x
R. lindleyi
. The left
half shows the appearance of the outside of the bud. The right half shows a cross section of the bud, with the rachis and two flowers in half tone. Four scales are shown on the outside of the bud. A calyx is shown next to the lower half of each flower. A bract is shown between the two flowers. Note the number of layers covering each flower near its base, and the number near its apex. Drawing by Pat Halligan |
Each flower has a characteristic orientation within the bud. The tangential (toward the outside) part of each flower faces the outside of the bud and receives more light than the axial (toward the axis of the bud) part of the flower. The bases of the dorsal (uppermost) and upper lateral lobes or petals of each flower face toward the axis of the bud. The bases of the lower lateral lobes of each flower face toward the outside of the bud.
The lobes themselves do not continue in this pattern, but are imbricated in a characteristic but variable pattern. The two upper lateral lobes generally occupy most of the outside of the upper half of the flower. The dorsal lobe usually occupies much of the axial side of the flower and typically shows only on the very tip of the tangential side of the flower. The two lower lateral lobes usually lie buried beneath the two upper lateral lobes.
When the bud opens and the flower emerges, the still closed flower is rather uniformly pink on the upper tangential portion and shades to greenish white toward the base and axial side. When the flowers begin to unfurl, white areas appear wherever previously covered areas become exposed by the separation of the lobes. As the flower expands, the reddish areas become progressively lighter and are separated by ever widening gulfs of white.
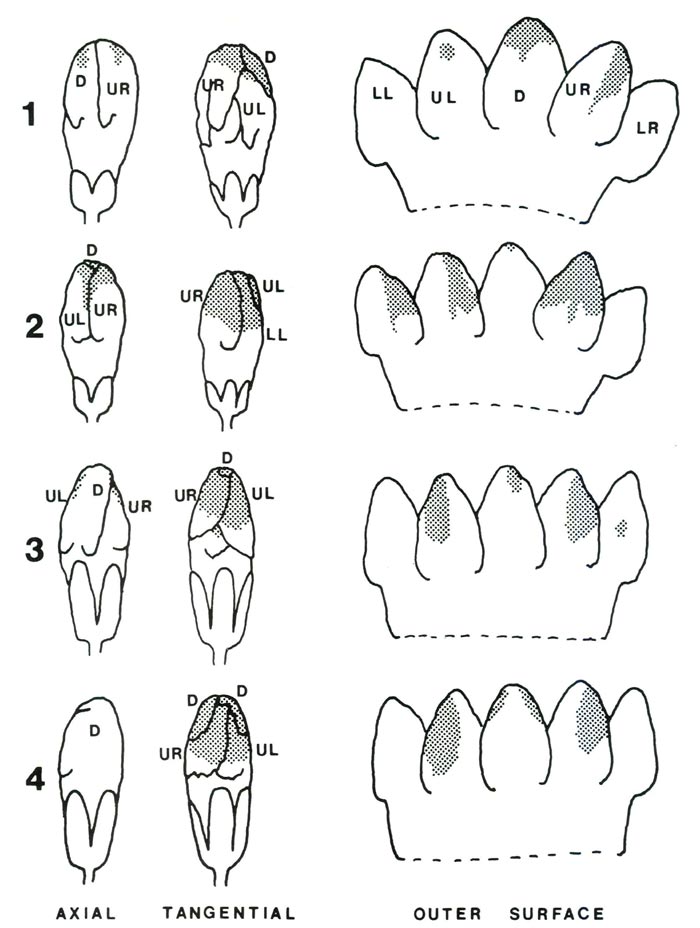
The pattern of imbrication varies, and the pigmentation patterning follows this pattern very closely, despite minor variations caused by folding and pleating of the corolla (Fig. 6). Those parts of the corolla exposed to the light in the bud end up as pink areas in the expanding flower.
|
|
Figure 6. Intact and dissected flowers taken from opening buds. Numbers 1 and 2 are
from 'Else Frye', numbers 3 and 4 are from R. nuttallii and R. lindleyi . The axial view is shown as though the viewer is standing at the center of the bud. The tangential view is shown as though the viewer is standing beside the bud. The dissected corollas are shown as though laid flat with the outside surface toward the viewer. Initials indicate the positions of each lobe: LL is lower left, UL is upper left, D is dorsal or uppermost, UR is upper right and LR is lower right. Drawings by Pat Halligan |
Testing Maddeniis
To test whether light impinging on the flowers within the bud has an effect on pigmentation patterning, I cut rectangular holes in the buds of
R. nuttallii
x
R. taggianum
(Fig. 7). In the bud pictured, no red pigment had formed yet when the hole was cut. The hole removed five layers of scales and calyx toward the base and three layers toward the apex, illustrating the fact that the covering of the bud is thinnest toward its apex.
Three days later, the corolla exposed by the hole had become uniformly red. After seven days (Fig. 8) the exposed portion of the corolla was no longer uniformly red, but was green at the base of the hole. After seventeen days the corolla seen through the hole was red only in the apical portion of the hole, and green in the basal two thirds.

|

|
|
|
Figure 7.
Expanding bud of R. nuttallii x R. taggianum with a rectangular hole cut in its side. This photo was taken immediately after the hole was cut. Photo by Pat Halligan |
Figure 8. The same bud as in Figure 7
after seven days. The corolla has turned red where exposed by the hole, except for the green band at the basal portion of the hole. This green band is newly exposed corolla which had been covered by bud scales basal to the hole. By the time this band had emerged into the open, the corolla was no longer turning red. Photo by Pat Halligan |
This change in color patterning can be explained by a sequence of pigment formation and corolla movement. Pigment was formed only for a short period after the hole was cut. Meanwhile the flowers were constantly moving apically relative to the hole as the bud expanded and opened. While pigment was being formed the entire hole was pigmented. After pigment ceased to be formed, the part of the corolla that was originally basal to the hole moved up into the hole and remained green. This experiment illustrates both the influence of light on the intensity of pigmentation, and the importance of the developmental stage on the pigmentation response to light.
This bud was allowed to develop into a fully formed truss. The flower that had been exposed by the hole ended up pink while the other flowers ended up white with only a trace of pink.
A partially expanded flower was dissected out of another bud in which a hole had earlier been cut. A pigmented area was found at the base of one of the lower lateral lobes in the area exposed by the hole. The atypical and asymmetric pigmentation pattern illustrates how manipulation of light can influence pigment patterning in the Maddeniis.
Since the pigments influenced by light were red or pink, I assume that the affected pigments are anthocyanins and flavonols. Of course, chemical studies' would be necessary to determine the precise nature of light influences upon pigmentation.
Patterning in Other Plants
Light induced pigmentation patterning apparently is not restricted to rhododendrons. I observed a pigmentation pattern in
Weigela hortensis
which was essentially the same as that in
R. campylogynum
x
R. spinuliferum
. The expanded but unopened corolla was uniformly red. When the lobes began to separate, white areas began to be exposed. When the corolla was fully opened, the outside was reddish except where the lobes had overlapped in the bud, where it was white. Inside, the corolla was almost white.
An unrelated plant, Gardenia imperialis , has been described as having a similar pattern: "corolla tube white inside, pink outside; corolla lobes white inside, pink and white flecked with deep red outside — the pink portion being the part exposed to the light when the flower is in "bud" (Eggerling and Dale, 1951). Once I had noticed light influenced pigmentation patterning in one flower, I started seeing it elsewhere.
Producing Hybrids With Uniform Color
It is important to recognize light induced pigmentation patterning when planning any breeding program. Crossing two plants with this pattern will almost surely produce progeny with the same pattern. This lies at the root of the failure to produce good uniform red flowers from
R. spinuliferum
. This species looks like a good uniform red parent until you examine the flower closely. The flower is lighter where the petals overlap and within the corolla. When this species is mated with other light dependent species, such as
R. racemosum
, the pattern comes through and the flowers of the hybrids are a disappointing blotchy pink. Taken to the second and third generations, the same disappointing pattern comes through, true to both parents. Uniform, intense flower color must be found elsewhere, in a species which is unaffected by light, such as
R. campylogynum
.
The same rationale applies to Maddenii hybrids. In order to produce hybrids with uniform intense color, the breeder must look elsewhere, to species which are unaffected by light. Unfortunately, the species not affected by light generally do not breed well with the Maddeniis. Just to prove that there is an exception to every rule, Bob Scott has produced an intensely colored, uniformly red Maddenii using species that shouldn't produce such a color pattern. This probably represents a mutation, which will most likely pass on to its progeny. Thus, from Bob Scott's red Maddenii will probably come the intensely colored Maddeniis we've been looking for.
Flowers with light dependent pigmentation patterning are notorious for their capricious nature. R. spinuliferum produces its reddest flowers when exposed to the cold, as do the various Maddeniis. Plants grown or forced in relatively warm greenhouses produce pale or even completely white flowers while those outdoors are highly colored. Although I did not test the effect of temperature on flower color, I hypothesize that the light dependent pigmentation forms best when cool temperatures are present. Both high light intensity and cool temperatures must be present for maximum production of pigment. Other plants, such as Gardenia imperialis , which grows in the steamy jungles of equitorial Africa, may not show this effect.
Reference
Eggerling, W. J., and I. R. Dale (1951)
The Indigenous Trees of the Uganda Protectorate.
The Government Printer, Entebbe, Uganda, Pg. 344.
Dr. Pat Halligan's hybridizing program was described in Smithsonian magazine (May 1986) as adapting Maddenii rhododendrons to a cooler environment. Previous articles for the ARS Journal include "Delayed Graft Incompatibility in Maddenii Rhododendrons" (Vol. 42:3, Fall 1987). Dr. Halligan, Seattle Chapter member, recently accepted the chairmanship of the ARS Plant Ratings Committee.
