JARS v63n3 - Fertility of Neopolyploid Rhododendron and Occurrence of Unreduced Gametes in Triploid Cultivars
Fertility of Neopolyploid Rhododendron and
Occurrence of Unreduced Gametes in Triploid Cultivars
Jeff R. Jones and Thomas G. Ranney
Department of Horticultural Science
Mountain Horticultural Crops Research and Extension Center
North Carolina State University
Mills
River, North Carolina
Introduction
Polyploidy, defined as an organism with three or more complete sets of chromosomes, is common in plants
and widely recognized as an important mechanism of adaptation and speciation (Ramsey and Schemske, 2002;
Soltis et al., 2003; Wendel, 2000). The prevalence of polyploidy in the genus
Rhododendron
has
previously been documented (Jones et al., 2007). In some cases, polyploid plants, including
Rhododendron
, can have desirable features including enhanced vigor, thicker leaves, and larger
flowers with thicker petals that persist longer (Kehr, 1996). Polyploidy can also have a profound
influence on reproductive biology, including fertility (Allard, 1960; Ramsey and Schemske, 1998). As
such, a greater understanding of reproductive behavior of polyploids is valuable to plant breeders.

|
The potential for utilizing polyploids in a breeding program is dependent upon fertility of specific taxa. The origin of polyploids can be a major factor in determining fertility. If a polyploid arises from somatic doubling in a meristem (spontaneous endoreduplication or chemical induction) or from the union of unreduced gametes from two closely related diploid parents, it will have four similar (homologous) versions of each chromosome and is often referred to as an autotetraploid (or polysomic tetraploid) (Ramsey and Schemske, 1998; Sanford, 1983). Autopolyploids may lack fertility due to the presence of multiple homologous chromosomes that can result in multivalent pairing and unequal segregation in meiosis (Ranney, 2006; Riesberg, 2001; Stebbins, 1950). Polyploids that result from somatic doubling in a hybrid or from unreduced gametes from different species are referred to as allopolyploids (or sometimes amphidiploids or disomic polyploids). Allopolyploids are often fertile due to nonrandom, disomic pairing between two distinct sets (pairing among chromosomes derived from each parental species, but not between species) of chromosomes during meiosis (Ramsey and Schemske, 2002; Ranney, 2006). In many cases, however, polyploids fall somewhere between an autopolyploid and an allopolyploid; where there is partial chromosome homology resulting in a combination of disomic and polysomic pairing; these polyploids are referred to as segmental allopolyploids. Surveying a broad range of plant species, Ramsey and Schemske (2002) noted that fertility varied considerably within both neoauto- and neoallopolyploids (newly formed polyploids), emphasizing the need to determine fertility on a plant-by-plant basis.
Triploids are often found to be highly infertile, if not sterile (Allard, 1960; Ramsey and Schemske, 1998). Infertility of triploids results from the fact that three sets of chromosomes cannot be divided evenly during meiosis yielding unequal segregation of the chromosomes often resulting in aneuploid gametes or meiotic failure. A number of triploid (2 n = 3x = 39) Rhododendron have been documented. De Schepper et al. (2001) confirmed triploidy in the azaleas 'Euratom', 'Euratom Orange'*, and 'Red Wing', and Jones et al. (2007) found 'Hallelujah', 'Taurus', 'White Ruffles'*, and 'Crimson Majesty'* to be triploids. However, little is known about the fertility of triploid Rhododendron .
Although triploids are often infertile, meiotic restitution can circumvent this limitation resulting in functional, unreduced gametes (Dweikat and Lyrene, 1988; Ramsey and Schemske, 1998). Triploids that produce unreduced gametes can be utilized as bridges for the development of tetraploids by crossing them back with diploids (Bretagnolle and Thompson, 1995; Ramsey and Schemske, 1998). Moreover, hexaploids may also be produced by intercrossing among triploids that produce viable unreduced gametes (Ehlenfeldt and Vorsa, 1993). Frequencies of unreduced gamete production are highly variable, may differ considerably among individuals, and vary with environmental factors such as temperature (Bretagnolle and Thompson, 1995; Dweikat and Thompson, 1988; Ramsey and Schemske, 1998). Unreduced pollen has previously been documented within the genus in diploid clones including the elepidote R. 'Hexe' (De Schepper et al. 2001), evergreen azaleas (Eeckhaut et al., 2006), and deciduous azaleas with hybrids involving R. x kosterianum and R. prinophyllum displaying a range of unreduced pollen production from 0 - 21% (Widrlechner et al., 1982). Circumstantial evidence also supports the role of unreduced gametes in the formation of various polyploid cultivars (Jones et al., 2007; Li, 1957; Widrlechner et al., 1982; Willingham, 1976). Unreduced gametes may also be more prevalent in triploids than in diploids (Dweikat and Lyrene, 1988; Veilleux, 1985). Examination of the fertility of specific clones is therefore necessary to determine the usefulness of incorporating triploids into a breeding program.
The pollen structure in Rhododendron and many Ericaceous species is typically a tetrad of grains that are tightly grouped at maturity (Copenhaver, 2005; Widrlechner et al., 1982). However, the pollen of unreduced gamete producers is often a mix of sporads containing tetrads, dyads, and monads (Ortiz et al., 1992). The dyad grain structure is representative of 2 n (the somatic chromosome complement) pollen that results from irregularities of the firs or second meiotic division (Bretagnolle and Thompson, 1995; Widrlechner et al., 1982). Unreduced pollen grains are also typically larger than reduced pollen grains (Bretagnolle and Thompson, 1995; Widrlechner et al., 1982). Pollen staining, using acetocarmine, can be used to assess male fertility and reflects the potential for pollen to germinate and contribute to fertilization (Dafni and Firmage, 2000). Acetocarmine stains nuclei within the cells and has been utilized previously in studies of Rhododendron pollen (Contreras et al., 2007; Sakai et al., 2004).
The objectives of this project were to: 1) evaluate the effect of increased ploidy level on pollen fertility of selected Rhododendron and 2) evaluate pollen fertility of naturally occurring triploids found in the genus.
| Table 1 . Pollen viability of polyploid Rhododendron and progenitor taxa. | ||||
| Taxa | Genotype | Ploidy Level | Viability (%) 1 | Contrast 2 |
| 'Nova Zembla' | 1 | 2x | 33.4 ∓ 1.8 | NS 3 |
| 'Super Nova' | 1 | 4x | 30.4 ∓ 2.5 | |
| 'Vulcan' | 2 | 2x | 48.0 ∓ 1.3 | NS |
| 'Vulcan' Tetraploid | 2 | 4x | 46.5 ∓ 6.4 | |
| 'Snowbird' | 3 | 4x | 63.8 ∓ 3.5 | P<0.0001 |
| 'Fragrant Star' | 3 | 8x | 16.4 ∓ 1.5 | |
| 'Fragrant Affinity'* | 4 | 2x | 1.5 ∓ 0.2 | P<0.0001 |
| 'Fragrant Affinity Tetra'* | 4 | 4x | 19.9 ∓ 2.3 | |
| fortunei | 5 | 2x | 47.4 ∓ 4.0 | P<0.0001 |
| fortunei Tetraploid | 5 | 4x | 7.7 ∓ 1.8 | |
| PJM Group | 6 | 2x | 31.6 ∓ 1.4 | P<0.0001 |
| 'Northern Starburst' | 6 | 4x | 47.0 ∓ 1.3 | |
| 'The Honourable Jean Marie de Montague' | 7 | 2x | 28.7 ∓ 2.8 | P<0.0001 |
| 'Briggs Red Star' | 7 | 4x | 11.0 ∓ 1.1 | |
| 'Weston's Aglo' | 8 | 2x | 19.6 ∓ 0.9 | P<0.0001 |
| 'Bubblegum' | 8 | 4x | 58.8 ∓ 3.2 | |
|
1
Values represent means ∓ SEM for 10 replications.
2 Contrast represents LSD, 0.05 mean separations between common (highlighted) genotypes of different ploidy levels. 3 NS = Not significant. |
||||
Materials and Methods
Comparing fertility between ploidy levels
. Newly formed polyploids and their progenitor taxa
were chosen to compare the fertilities between ploidy levels within the same genotype. The ploidy levels
of all plants were determined by Jones et al. (2007) (Table 1). The tetraploid R. 'Supernova' and the
octoploid R. 'Fragrant Star' were chemically induced at Briggs Nursery, Olympia, Wash., from R. 'Nova
Zembla' and R. 'Snowbird', respectively (Dan Meier, Olympia, Wash., per. comm.). Rhododendron 'Vulcan'
(tetraploid form) is a 2x + 4x mixaploid (Jones et al., 2007) and the result of natural somatic
endoreduplication in a shoot of R. 'Vulcan' (Harold Greer, Eugene, Ore., per. comm.). Rhododendron
'Briggs Red Star' is also a 2x + 4x mixaploid (Jones et al., 2007) developed at Briggs Nursery through
the chemical induction of R. 'The Honourable Jean Marie de Montague'. Both mixaploids appear to be
tetraploid in the LII histogenic layers since they breed as tetraploids (Dr. Thomas G. Ranney,
N.C. State University, NC, per. comm.) and gametogenesis occurs within the LII layer (Pratt, 1983;
Ranney, 2006). Rhododendron 'Fragrant Affinity Tetra'* was chemically induced from R. 'Fragrant
Affinity'* (Contreras et al., 2007). Dr. Max Byrkit chemically induced
R. fortunei
to produce
the tetraploid form (Kehr, 1996). Rhododendron 'Northern Starburst' and R. 'Bubblegum' are both
chemically induced tetraploids of R. PJM Group and R. 'Weston's Aglo', respectively, developed at
Briggs Nursery (Dan Meier, Olympia, Wash., per. comm.).
Pollen viability was determined using staining procedures similar to those described by Contreras et al. (2007) and Olsen et al. (2006). All pollen was collected at anthesis from plants at the Mountain Horticultural Crops Research Station, dried at ~21°C for 24 hrs, and stored at -25°C until testing. Pollen was placed on glass microscope slides, and the grains were stained with 1% acetocarmine (w/v) for 15 minutes. Pollen grains that stained a distinct red-pink color were scored as viable (Dafni and Firmage, 2000; Ramsey and Schemske, 2002). The tetrad nature of Rhododendron pollen required each individual grain in the tetrad to be analyzed. Each tetrad has the potential to contain four viable grains. The experimental design was a randomized complete block with ten replicates blocked over time. A minimum of 50 tetrads were randomly selected and analyzed per replicate. Pollen was observed at 300x using a light microscope (Nikon Eclipse 80i, Nikon, Melville, NY). Pollen viability percentages were calculated and the data were subjected to analysis of variance and pairwise means comparisons between ploidy levels for a given genotype (LSMEANS option, PROC GLM; SAS version 8.02, SAS Institute., Cary, N.C.; SAS Institute, 1988).
Triploid fertility . The cultivars R. 'Hallelujah', R. 'Red Wing', and R. 'Taurus' were confirmed as triploids by Jones et al. (2007). Recent analysis also determined that the azaleodendrons (crosses of an unnamed Ilam azalea by the evergreen R. 'Catalgla' provided by Dr. Steven Krebs at the Holden Arboretum, Kirtland, Ohio) 94-28/2, 94-28/3, 94-28/7, and 94-28/14 were also triploids (Krebs, S.L., unpub. material). Pollen was collected at anthesis, and the frequency of viable, unreduced gametes in the triploid taxa was determined using pollen staining as described above and observed under a light microscope at 300x. The experimental design was a randomized complete block with 5 replicates blocked over time. At least 50 sporads (tetrads, dyads, or monads) were randomly selected and analyzed per replicate. Pollen was considered viable and unreduced if there was a well-stained monad or dyad and the pollen diameter was visibly larger (>120%) than normal (Shoemaker-Megalos and Ballington, 1988). The frequency of unreduced gametes was determined using the equation from Ortiz et al. (1992):
Unreduced pollen frequency = [(2 x # of dyads) + (# of monads)] / (# of total grains). Data were then subjected to analysis of variance and means compared using least significant differences (LSD) (PROC GLM; SAS version 8.02, SAS Institute., Cary, N.C.; SAS Institute, 1988).
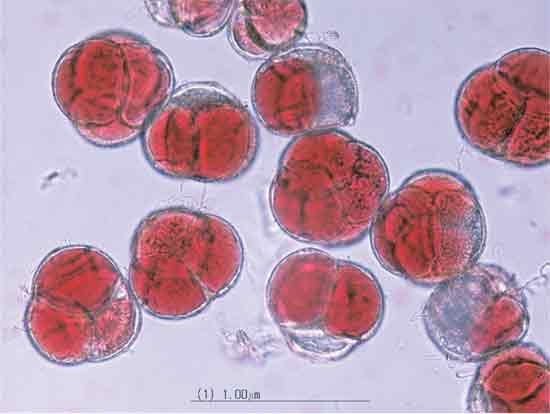
|
|
Figure 1. Photomicrograph of
tetrad pollen grains of
the tetraploid Rhododendron 'Northern Starburst'. |
Results and Discussion
Comparing fertility between ploidy levels. Pollen grains were readily apparent as being stained or
unstained (Figure 1). Pollen viabilities for all taxa ranged from 1.5 - 63.8% (Table 1). There
was a significant effect of ploidy level (P<0.001), genotype (P<0.0001), and a ploidy-genotype
interaction (P<0.0001) on fertility. The significant interaction indicated the effect of polyploidy on
fertility depended on genotype.
In the genotypes, R. fortunei , 'The Honourable Jean Marie de Montague', and 'Snowbird', the increased ploidy level reduced fertility. The tetraploid form of R. fortunei is an induced autotetraploid and its reduced fertility most likely results from problems associated with polysomic chromosome pairing commonly found in autotetraploids. The diploid R. 'The Honourable Jean Marie de Montague' is a hybrid of unknown parentage believed to involve R. griffithianum . The reduced fertility of the tetraploid R. 'Briggs Red Star' suggests that it functions more like an autotetraploid or segmental allopolyploid and the increased ploidy level compromised gamete formation. Rhododendron 'Snowbird' is a naturally occurring tetraploid believed to be a hybrid between R. atlanticum and R. canescens (Galle, 1987), most likely the result of a normally reduced gamete from R. atlanticum combined with an unreduced gamete from R. canescens . The induced octoploid R. 'Fragrant Star' would thus be considered an autoallopolyploid, that contains four homologous chromosomes from each species, which may still result in polysomic chromosome pairing leading to reduced fertility.
In the genotypes, R. PJM Group, 'Weston's Aglo', and 'Fragrant Affinity'*, the increased ploidy level significantly increased fertility. Both R. PJM Group and 'Weston's Aglo' are interspecific hybrids between R. minus x R. dauricum . The induced polyploids of these taxa, R. 'Northern Starburst' and 'Bubblegum', represent allopolyploids, and the increased fertility most likely results from restoration of chromosome homology and improved disomic pairing. The increased fertility of R. 'Fragrant Affinity Tetra'* over 'Fragrant Affinity'* represents a similar case as previously described by Contreras et al. (2007).
For the remaining genotypes, R. 'Nova Zembla' and R. 'Vulcan', increased ploidy level had no apparent effect on fertility. Both of these genotypes are complex hybrids and the induced tetraploids most likely function as segmental allopolyploids with moderate to high fertility, regardless of ploidy level.
These results demonstrate that fertility of polyploid Rhododendron can be highly variable and that induced polyploidy may either enhance or compromise fertility. The effect of polyploidy on fertility most likely results from the level of homology among the chromosome sets and subsequent impacts on chromosome pairing during meiosis. Information on the fertility of specific cultivars will assist breeders in utilizing these plants in future breeding programs.
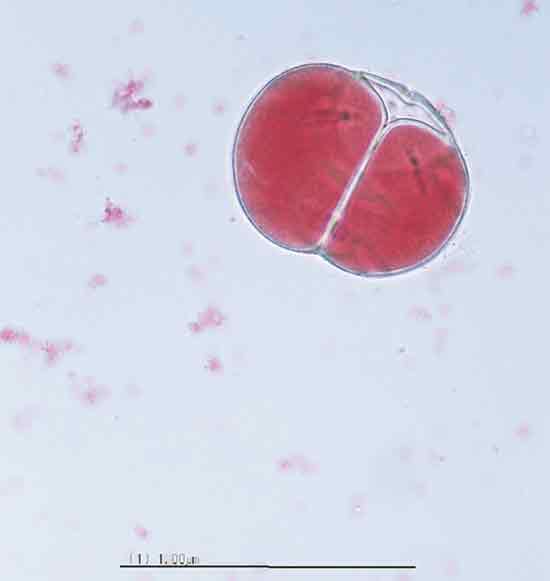
|
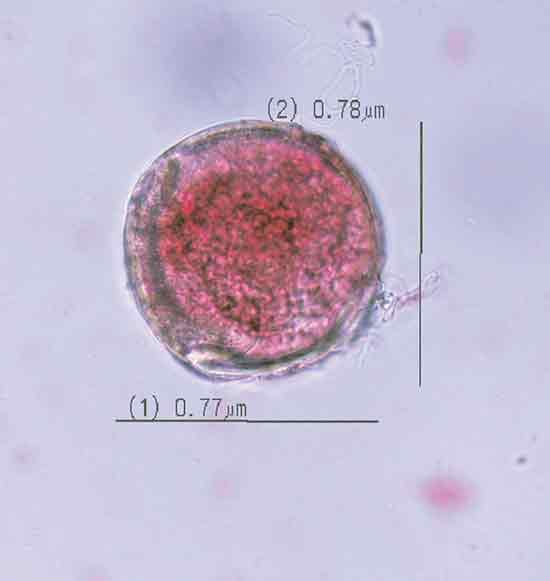
|
|
|
Figure 2. Photomicrograph of
dyad pollen grain
of the triploid Rhododendron 'Red Wing'. |
Figure 3. Photomicrograph of
monad pollen grain of the triploid
Rhododendron 'Hallelujah' illustrating size dimensions. |
Triploid fertility . Viable dyad and monad grains (Figures 2 and 3) were observed in pollen samples from triploid taxa, ranging from 0.2 to 5.3% (Table 2), indicating the presence of unreduced pollen. The increased size of unreduced pollen diameter was also clearly evident (Figure 3). As in other triploid Ericaeous taxa (Ehelnfeldt and Vorsa, 1993; Vorsa and Ballington, 1991), the small percentage of viable stained pollen was largely limited to monad or dyad grains. There was a significant effect (P<0.0001) of taxa on the percentage of unreduced pollen. Rhododendron 'Red Wing' had the highest percentage of unreduced pollen at 5.3%, followed by R. 'Hallelujah' at 2.9%, while the remaining taxa were similar with less than 1.1% unreduced gametes. In studies of Vaccinium spp., also in the Ericaceae, significant differences in frequencies of unreduced pollen among taxa have also been reported (Ortiz et al., 1992; Shoemaker-Megalos and Ballington, 1988).
| Table 2 . Percent unreduced gametes in selected triploid Rhododendron taxa. | |
| Taxa | Viable Unreduced Gametes (%) 1 |
| 'Hallelujah' | 2.87 ∓ 0.55 B |
| 'Red Wing' | 5.31 ∓ 0.81 A |
| 'Taurus' | 1.09 ∓ 0.13 C |
| 'White Ruffles' | 0.65 ∓ 0.19 C |
| Azaleodendron 94-28/2 | 0.45 ∓ 0.19 C |
| Azaleodendron 94-28/3 | 0.98 ∓ 0.27 C |
| Azaleodendron 94-28/7 | 0.60 ∓ 0.23 C |
| Azaleodendron 94-28/14 | 0.23 ∓ 0.01 C |
| 1 Values represent means ∓ SEM. Means followed by a different letter are significantly different at P<0.05. | |
Dyad pollen grains are typically expected to be 2
n
and can result from a variety of meiotic
irregularities including first or second division restitution (Bretagnolle and Thompson, 1995;
Islam and Shepherd, 1980; Widrlechner, 1982). Monad pollen grains could result from different
pathways including double restitution (4
n
) or the combination of a restitution event and
abnormal nuclear fusion of individual grains producing a 2
n
monad (Bretagnolle and Thompson,
1995; Veilleux, 1985).
The production of unreduced pollen by triploid taxa indicates the potential for utilizing certain
taxa in breeding programs. The greater frequency of unreduced pollen found in R. 'Red Wing' and
'Hallelujah' may allow for successful hybridizations given adequate numbers of pollinations.
Additional breeding studies with triploid parents would help elucidate the level of functional
unreduced pollen and the presence of unreduced female gametes (Ehlenfeldt and Vorsa, 1993).
Overall, the influence of polyploidy on fertility in Rhododendron is highly variable and appears to be influenced by the ploidy level, degree of homology among chromosomes, and in the case of triploids, the frequency of unreduced gamete formation. A greater understanding of fertility mechanisms in polyploid Rhododendron and information on fertility of specific clones will better allow breeders to incorporate and utilize polyploids in plant improvement programs.
* = name of plant is unregistered
Literature Cited
Allard, R.W. 1960.
Principles of Plant Breeding
. Wiley Pub., New York, NY: 264pp.
Bretagnolle, F. and J.D. Thompson. 1995. Tamsley review no. 78. Gametes with the somatic
chromosome number: mechanisms of their formation and role in the evolution of autopolyploid
plants.
New Phytol
. 129: 1-22.
Contreras, R.N., T.G. Ranney, and S.P. Tallury. 2007. Reproductive behavior of diploid and
allotetraploid
Rhododendron
L. 'Fragrant Affinity'.
HortScience
42: 31-34.
Copenhaver, G.P. 2005. A compendium of plant species producing pollen tetrads.
J. North Carolina Acad. Sci
. 121: 17-35.
Dafni, A. and D. Firmage. 2000. Pollen viability and longevity: practical, ecological and evolutionary
implications.
Plant Syst. Evol
. 222: 113-132.
De Schepper, S., L. Leus, M. Mertens, E. Van Bockstaele, and M. De Loose. 2001. Flow cytometric
analysis of ploidy in
Rhododendron
(subgenus
Tsutsusi
).
HortScience
36: 125-127.
Dweikat, I.M. and P.M. Lyrene. 1988. Production and viability of unreduced gametes in triploid
interspecific blueberry hybrids.
Theor. Appl. Genet
. 76: 555-559.
Eeckhaut, T.G., J. Van Huylenbroeck, S. De Schepper, and M.C. Van Labeke. 2006. Breeding for
polyploidy in Belgian azalea (
Rhododendron simsii
hybrids). Proc. XXIInd Intl. Eucarpia
Symp. on Breeding for Beauty,
Acta Hort
. 714: 113-118.
Ehlenfeldt, M.K. and N. Vorsa. 1993. The generation, evaluation and utilization of hexaploid
progeny from 3x x 3x crosses of highbush blueberry: germplasm transfer and 2n gametes.
Acta Hort
. 346: 95-109.
Galle, F.C. 1987.
Azaleas: Revised and enlarged edition
. Timber Press, Portland, OR: 519pp.
Islam, A.K.M.R. and K.W. Shepherd. 1980. Meiotic restitution in wheat-barley hybrids.
Chromosoma
79: 363-372.
Jones, J.R., T.G. Ranney, N.P. Lynch, and S.L. Krebs. 2007. Ploidy levels and
genome sizes of diverse species, hybrids, and cultivars of
Rhododendron
.
J. Amer. Rhododendron Soc
. 61: 220-227.
Kehr, A.E. 1996. Polyploids in rhododendron breeding.
J. Amer. Rhododendron Soc
. 50: 215-217.
Li, H. 1957. Chromosome studies in the azaleas of eastern North America.
Amer. J. Bot
. 44:
8-14.
Olsen, R.T., T.G. Ranney, and Z. Viloria. 2006. Reproductive behavior of induced allotetraploid
x Chiltalpa and in vitro embryo culture of polyploidy progeny.
J. Amer. Soc. Hort. Sci
. 131:
716-724.
Ortiz, R., N. Vorsa, L.P. Bruederle, and T. Laverty. 1992. Occurrence of unreduced pollen in
diploid blueberry species,
Vaccinium
sect.
Cyanococcus
.
Theor. Appl. Genet
. 85: 55-60.
Pratt, C. 1983. Somatic selection and chimeras. In: J.N. Moore and J. Janick (eds.).
Methods in fruit breeding
. Purdue Unv. Press, West Lafayette, Ind.: 172-185.
Ramsey, J. and D.W. Schemske. 1998. Pathways, mechanisms, and rates of polyploidy formation in
flowering plants.
Annu. Rev. Ecol. Syst
. 29: 467-501.
Ramsey, J. and D.W. Schemske. 2002. Neopolyploidy in flowering plants.
Annu. Rev. Ecol. Syst
.
33: 589-639.
Ranney, T.G. 2006. Polyploidy: From evolution to new plant development.
Proc. Intern. Plant
Propagators' Soc
. 56: 137-142.
Riesberg, L.H. 2001. Polyploid evolution: keeping the peace at genomic reunions.
Curr. Biol
.
11: R925-R928.
Sanford, J.C. 1983. Ploidy manipulations. In: J.N. Moore and J. Janick (eds.).
Methods in fruit
breeding
. Purdue Univ. Press, West Lafayette, Ind.: 100-123.
SAS Institute Inc. 1988.
SAS/STAT user's guide
, release 6.03 edition. SAS Institute Inc.,
Cary, NC.
Sakai, K., I. Miyajima, K. Ureshino, Y. Ozaki, and H. Okubo. 2004. Oryzalin-induced
allotetraploids of an intersubgeneric hybrid between evergreen and deciduous azaleas. J
. Fac.
Agri
. Kyushu Univ. 49: 293-299.
Shoemaker-Megalos, B. and J.R. Ballington. 1988. Unreduced pollen frequencies versus hybrid
production in diploid - tetraploid
Vaccinium
crosses.
Euphytica
39: 271-278.
Soltis, D.E., P.S. Soltis, and J.A. Tate. 2003. Advances in the study of polyploidy
since plant speciation.
New Phytol
. 161: 173-191.
Stebbins, G.L., Jr. 1950.
Variation and evolution in plants
. Columbia Univ. Press, New York,
NY: 57pp.
Veilleux, R. 1985. Diploid and polyploidy gametes in crop plants: mechanisms of formation and
utilization in plant breeding.
Plant Breeding Reviews
3: 253-288.
Vorsa, N. and J.R. Ballington. 1991. Fertility of triploid highbush blueberry.
J. Amer. Soc.
Hort. Sci
. 116: 336-341.
Wendel, J.F. 2000. Genome evolution in polyploids.
Plant Mol. Bio
. 42: 225-249.
Widrlechner, M., H. Pellett, and P. Ascher. 1982. Unreduced gametes in azalea hybrids: a
possible breeding method for using promising azaleas of low fertility.
J. Amer. Rhododendron
Soc
. 36: 98-100.
Willingham, F.F., Jr. 1976. Variation and phenological forms in
Rhododendron calendulaceum
(Michx.) Torrey (Ericaceae).
Castanea
41: 215-223.
Acknowledgments
Much appreciation is given to Mr. Tom Eaker, Mr. Joel Mowrey, Mr. Nathan Lynch, and the staff
of the Mountain Horticultural Crops Research Station for their superb technical assistance
and to Dr. Stephen Krebs at the Holden Arboretum for provided plant material. Partial
funding for this project was provided by the Research Foundation of the American
Rhododendron Society.

Jeff R. Jones was a Master of Science student.
Thomas G. Ranney is a professor of Horticultural Science and member of the Southeastern Chapter.
